Taxonomy
Class: AmphibiaOrder: AnuraFamily: RanidaeSynonym: Rana clamitans
Taxonomic Comments: Frost et al. (2006) placed species in North American that were formerly in the very large genus Rana into a separate genus, Lithobates , to distinguish them from a large and predominantly Eurasian genus Rana (sensu stricto). There have been numerous arguments put forth for well over a decade about whether these species should be placed back into Rana or retained as Lithobates , with some supporting placing Lithobates as a subgroup within Rana and others supporting the recognition of both genera. There has been no clear resolution of the issue, and both Lithobates and Rana continue to be widely used in recent published literature on North American species. Here, we follow the recommendations of the Society for the Study of Amphibians and Reptiles' Standard English Names Committee and use Lithobates for North American representatives of this group. Rana clamitans melanota ) and the Bronze Frog (Rana c. clamitans ). These are no longer recognized and are treated as one of several color phases of a single species, the Green Frog (Lithobates clamitans ).Species Comments:
Identification
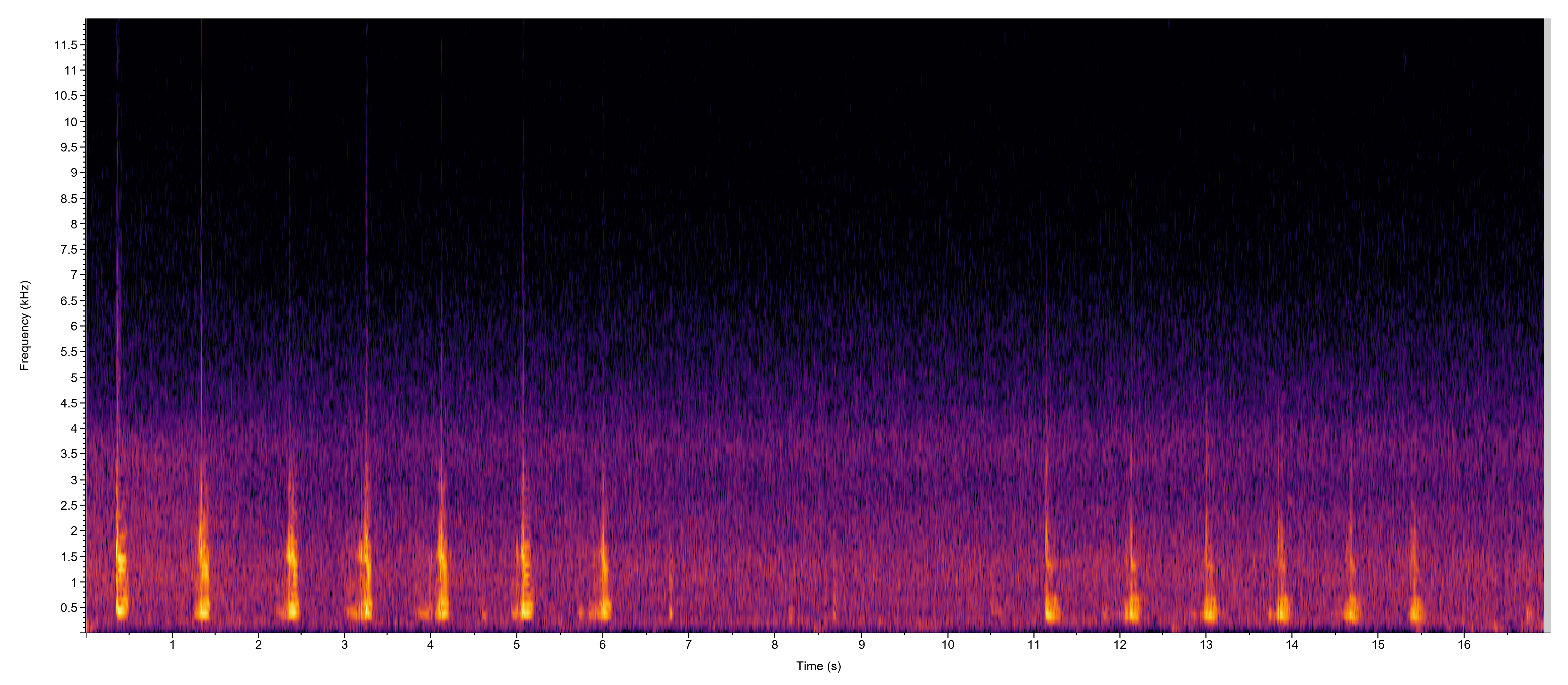
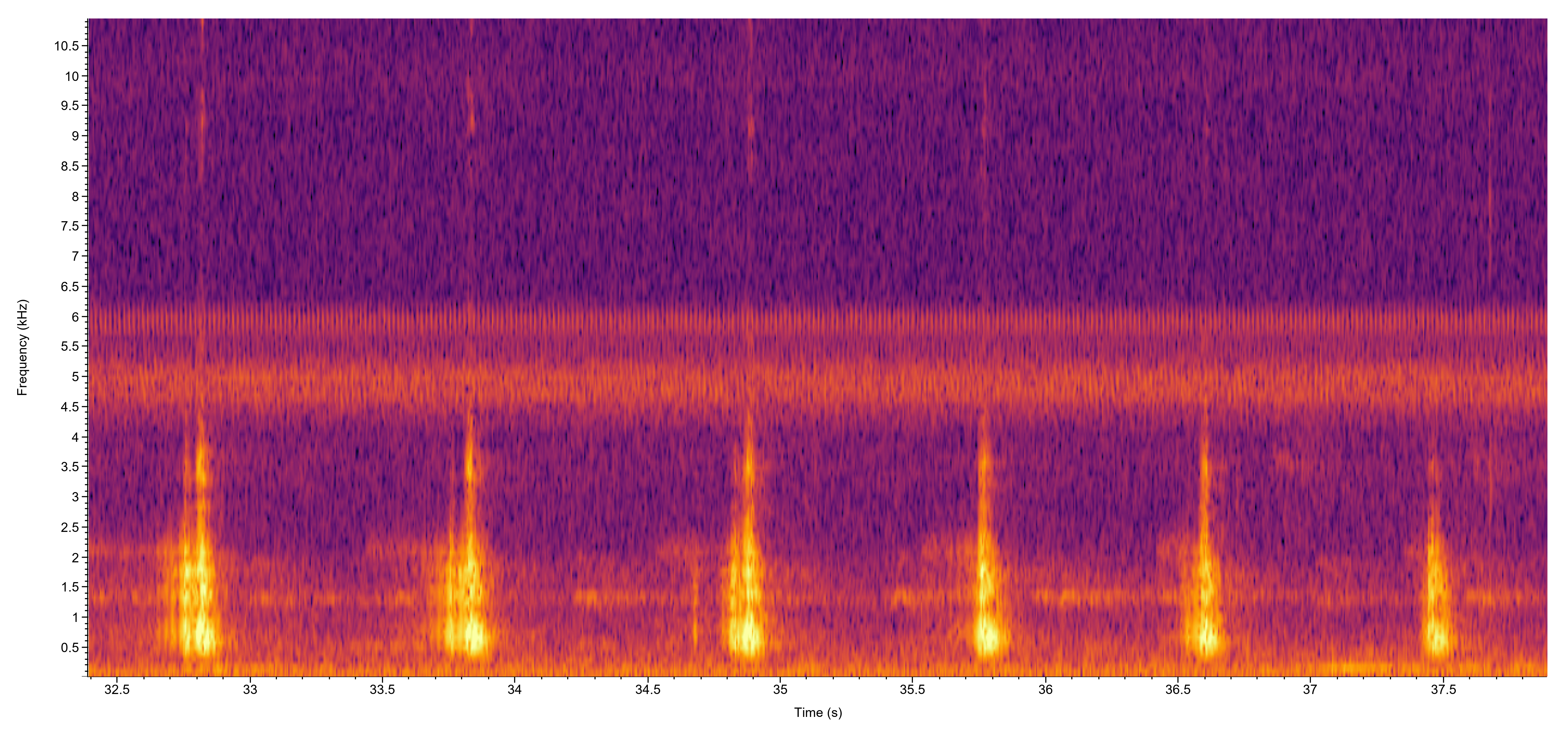
Description: The adults are medium-sized frogs that exhibit substantial geographic variation in coloration and body size. Two color phases that were previously recognized as subspecies occur throughout the range. These are a green form that is more northern and commonly referred to as the Northern Green Frog, and a southern bronze form commonly referred to as the Bronze Frog. In addition to these regional groups, some northern populations in scattered populations from the Great Lakes region to the Northeast are extremely dark olive to black and are commonly referred to as the Black Frog (Dodd 2013, Mecham 1954). 'Green Frog' is the standardized common name that now applies to all populations of this species, regardless of the particular color form. North Carolina has both the green and bronze forms, as well as numerous intermediate types, with bronze individuals predominating in the southeastern part of the state (Beane et al. 2010). Many individuals in North Carolina with greenish heads have the green coloration restricted to the sides of the head. Lithobates clamitans is best identified by a dorsolateral fold that extends on each side from behind the eye down the back before fragmenting about midway or so. A secondary fold branches off of the dorsolateral fold and extends around the tympanum. Populations in the northern part of the range can vary from green to olive green or brown above. Their skin tends to be somewhat rugose and the head is usually bright green. The dorsum, legs, and sides are often flecked with black spots, and the legs lack black bars above. The side of the body tends to have a dark, wavy, indistinct band that phases into darker mottling near the venter below. The center of the brownish tympanum sometimes has a white or yellow center and the throat is often darkly mottled to varying degrees. The belly is grayish white to whitish with varying levels of darker mottling. When present, the darker mottling is usually confined to the margins and lower sides, but individuals are occasionally seen that are more boldly marked with dusky reticulations across the entire belly (Dodd 2013). Vocalizations: The males produce several types of calls, with the advertisement call the most well recognized one. It may consists of a single note that resembles a deep "c'tung" or a rapid sequence that has 2-7 notes (e.g., "c'tung - c'tung - c'tung - c'tung." The larger males have deeper voices than the younger ones, and the number of notes per call typically increases with overall chorus activity during a night (Wells 1978). Bee et al. (2008) analyzed individual advertisement calls of males and found that they were distinctive. The males are likely able to discriminate between strangers and adjacently territorial holders based on individual variation in advertisement calls. Technical Reference: Dodd (2013)Online Photos: Google iNaturalist Observation Methods: Green frogs are most easily observed by walking the margins of wetlands or searching roads on rainy nights. Their distinctive advertisement call is readily heard around breeding sites during the summer months.
Download Video:
"MP4"
AmphibiaWeb Account
Distribution in North Carolina
Distribution Comments: The Green Frog is widely distributed in eastern North America, including much of extreme southern Canada from southeastern Manitoba eastward to the Canadian Maritimes and St. Lawrence River Valley. The range includes much of the eastern US, with the southern limit in central Florida, the Gulf States, and eastern Texas. The western limit includes eastern Texas, eastern Oklahoma, central and southeastern Missouri, eastern Iowa, and eastern Minnesota. Populations have been introduced in western North America, including British Columbia, Washington, Arizona, Iowa, and Montana (Dodd 2013). This species occurs statewide in North Carolina where it has been found in almost every county in the state. Distribution Reference: Dodd (2013)County Map: Clicking on a county returns the records for the species in that county.
GBIF Global Distribution
Key Habitat Requirements
Habitat: Lithobates clamitans is a semiaquatic species that generally stays close to bodies of water that are used for foraging, breeding, and overwintering. The juveniles and adults use all sorts of aquatic habitats, including both standing water, springs, and the sluggish portions of streams. The length of the larval period is variable depending on latitude and local site conditions. Many populations require a year or more to metamorphose and semipermanent or permanent habitats are requires to support local populations. The larvae are somewhat unpalatable to fish -- and have behavioral defenses also -- but in many areas they do best in either fish-free permanent sites or sites with well-developed, vegetated shallows that provide cover and protection for the larvae and adults (Richardson 2001, Werner and McPeek 1994). Environmental and Physiological Tolerances: The Green Frog appears to be moderately tolerant of acidic waters. It is generally restricted to breeding sites with a pH above 4.0 or higher, with local abundance being positively correlated with pH (Dodd 2013).Biotic Relationships: The larvae typically live in semipermanent or permanent bodies of water that have numerous aquatic predators. Fish will readily eat the ovarian eggs, suggesting that they are palatable to predators (Licht 1969). However, the jelly envelope may offer a defense against fish and other predators (Smith et al. 2019). There is limited evidence that ostracods may also eat the eggs under natural conditions (Gray et al. 2010). Some of the documented predators on the larvae include predaceous diving beetles, water beetles (Hydrophylidae), whirly gig beetles, giant water bugs, water scorpions, backswimmers, dragonfly larvae, crayfishes, turtles, and fishes (Dodd 2013, Smith et al. 2019). Ambystoma tigrinum larvae. L. catesbeianus , L. clamitans ), turtles (e.g., Chelydra serpentina ), snakes (Agkistrodon ; Nerodia ; Thamnophis ), crows, herons, hawks, and presumably mammals such as mink, otters, and raccoons (Dodd 2013). The juveniles and adults appear to minimize risk primarily via camouflage and remaining immobile, then leaping into the water if approached. A startle call is typically emitted when leaping that helps to disorient predators. Individuals may either crouch when approached by a snake -- which reduces their detectability -- or inflate their bodies to look more intimidating (Dodd 2013, Heinen and Hammond 1997, Marchisin and Anderson 1978).
Life History and Autecology
Breeding and Courtship: The adults breed during the warmer months of the year, but patterns are geographically variable. Dodd (2013) and Meshaka et al. (2009a) have comprehensive summaries of geographic patterns throughout the range. The calling and breeding season tends to be more prolonged in southern and coastal populations relative to those in northern or mountainous areas (e.g., Berven et al. 1979). The males begin calling with the arrival of warm weather in late winter, spring or early summer and may continue through August or September depending on the location and site conditions. Actual breeding and egg laying tends to begins a few weeks after the males begin calling and is more concentrated during the first few months of the breeding season. Reproductive Mode: When mating, each female produces a single, irregular, surface film (monolayer) of eggs that are deposited in shallow, vegetated areas of the breeding site. The surface film is typically < 30 cm in diameter and may either float freely or be anchored to vegetation (Dodd 2013). It is similar to that of the American Bullfrog, but smaller and contains far fewer eggs. The freshly laid eggs are black above and whitish below, and each is surrounded by two jelly envelopes that may fuse with those of neighboring eggs with time. The eggs vary from 1.2–2.1 mm in diameter, and the outer capsule is around 5–6 mm in diameter (Dodd 2013, Meshaka and Hughes 2014). Hatching normally occurs in three to five days.Aquatic Life History: The hatchlings and small larvae disperse from the ovipositing sites and do not form schools or other active social groups. They are active swimmers and feeders relative to most other ranids and often concentrate in warm, vegetated shallows during the warmer periods of each day to feed. Warkentin (1992) observed that the older tadpoles often moved from shallow water to deeper water at night, perhaps as a way to avoid shoreline predators. Terrestrial Life History: The tadpoles metamorphose during the warmer months of the year (April-October), and the young metamorphs spend only a short time at the breeding sites before dispersing into the surrounding country. Some may remain in the vicinity of a breeding site, but the great majority move away (Dodd 2013). Almost all of several hundred frogs that were tracked by Schroeder (1976) in Virginia left the breeding pond within a month. They moved away in all directions and used suitable habitat such as moist stream drainages and other wetlands along the way. Most frogs were recaptured from 183–448 m away, but eight were recaptured in an adjoining bog 2.5 km away, and two others 4.8 km away. Metamorphs at Martof’s (1953a) Michigan site only stayed for a week or so after transforming before moving to nearby streams. L. clamitans reaches sexual maturity at a relative small size (e.g., 40 mm for males and 43 mm for females in southern Louisiana) and can become sexually mature in as little as 3-4 months after metamorphosing. Both sexes breed within a year or so after metamorphosing. Similar patterns were observed in specimens from northern Florida (Meshaka and Marshall 2012). Acris gryllus ) and Spring Peeper (Pseudacris crucifer ). There are three records of a Green Frog eating a hatchling Spotted Turtle (Clemmys guttata ), a Meadow Jumping Mouse (Zapus hudsonius ), and a Big Brown Bat (Eptesicus fuscus ).
General Ecology
Population Ecology: Green Frog populations are probably not organized as metapopulations either locally or regionally given their ability to disperse long distances and their lack of strong habitat fidelity (Smith and Green 2005). Populations that use local breeding sites appear to be highly interconnected in many regions, and local ponds in some areas have turnover rates with recolonization occurring rapidly (Hecnar and M’Closkey 1996a, Skelly et al. 2003).Community Ecology: Lithobates clamitans often shares breeding ponds with one or more large ranids and may interact via both competition and predation. The larvae often coexist with L. americanus , and studies by Werner (1994) suggests that bullfrog tadpoles may be superior competitors overall. L. clamitans , for example, can displace male L. virgatipes from calling sites (Given 1990). Competition for food with ranids such as the Northern Leopard Frog, American Bullfrog, and Mink Frog is possible due to the generalist diets of these species (Stewart and Sandison 1972). These species often spatially segregate to varying degrees where they share habitats, which may lessen competitive interactions (McAlpine and Dilworth 1989). The American Bullfrog often preys on smaller ranids, including the Green Frog. Hecnar and M’Closkey (1997a) found that Green Frog numbers increased markedly at a site in Ontario after bullfrogs went extinct, presumably due to release from predation and/or competition.
Adverse Environmental Impacts
Status in North Carolina
NHP State Rank: S5Global Rank: G5Status Comments: This is one of the most common frogs in North Carolina and it tolerates habitat disturbance and alteration better than most species. Populations often occur on timbered landscapes so long as breeding sites are present, and they readily use farm ponds and lakes as breeding sites which helps compensate for the loss of natural habitats. Populations in North Carolina appear to be stable and show no evidence of significant declines.
Photo Gallery for Lithobates clamitans - Green Frog 22 photos are shown. Recorded by: Khaos Recorded by: Khaos Recorded by: Emily Stanley Recorded by: B. Bockhahn, T. Abee Recorded by: Steve Hall, Carol Tingley, Tom Howard, David George, Pat Coin, Jeff Niznik, Recorded by: Kate Hammett Recorded by: R. Miller Recorded by: David George, Jeff Niznik Recorded by: K. Bischof Recorded by: Travis McLain Recorded by: Steve Hall Recorded by: Jim Petranka Recorded by: Steve Hall and Bo Sullivan Recorded by: Steve Hall and Bo Sullivan Recorded by: L. Osteen Recorded by: tom ward Recorded by: tom ward Recorded by: tom ward Recorded by: John Petranka Recorded by: K. Long Recorded by: Steve Hall Recorded by: Steve Hall

 »
»