Taxonomy
Class: AmphibiaOrder: AnuraFamily: RanidaeSynonym: Rana sphenocephala
Taxonomic Comments: Frost et al. (2006) placed species in North American that were formerly in the very large genus Rana into a separate genus, Lithobates , to distinguish them from a large and predominantly Eurasian genus Rana (sensu stricto). There have been numerous arguments put forth for well over a decade about whether these species should be placed back into Rana or retained as Lithobates , with some supporting placing Lithobates as a subgroup within Rana and others supporting the recognition of both genera. There has been no clear resolution of the issue, and both Lithobates and Rana continue to be widely used in recent published literature on North American species. Here, we follow the recommendations of the Society for the Study of Amphibians and Reptiles' Standard English Names Committee and use Lithobates for North American representatives of this group. Barrow et al. (2018) analyzed mtDNA variation in populations in the Southeast and found evidence of two deeply divergent eastern and western mtDNA clades that meet in central Alabama. In contrast, analyses of nuclear genes resulted in poorly resolved gene trees. Species Comments:
Identification
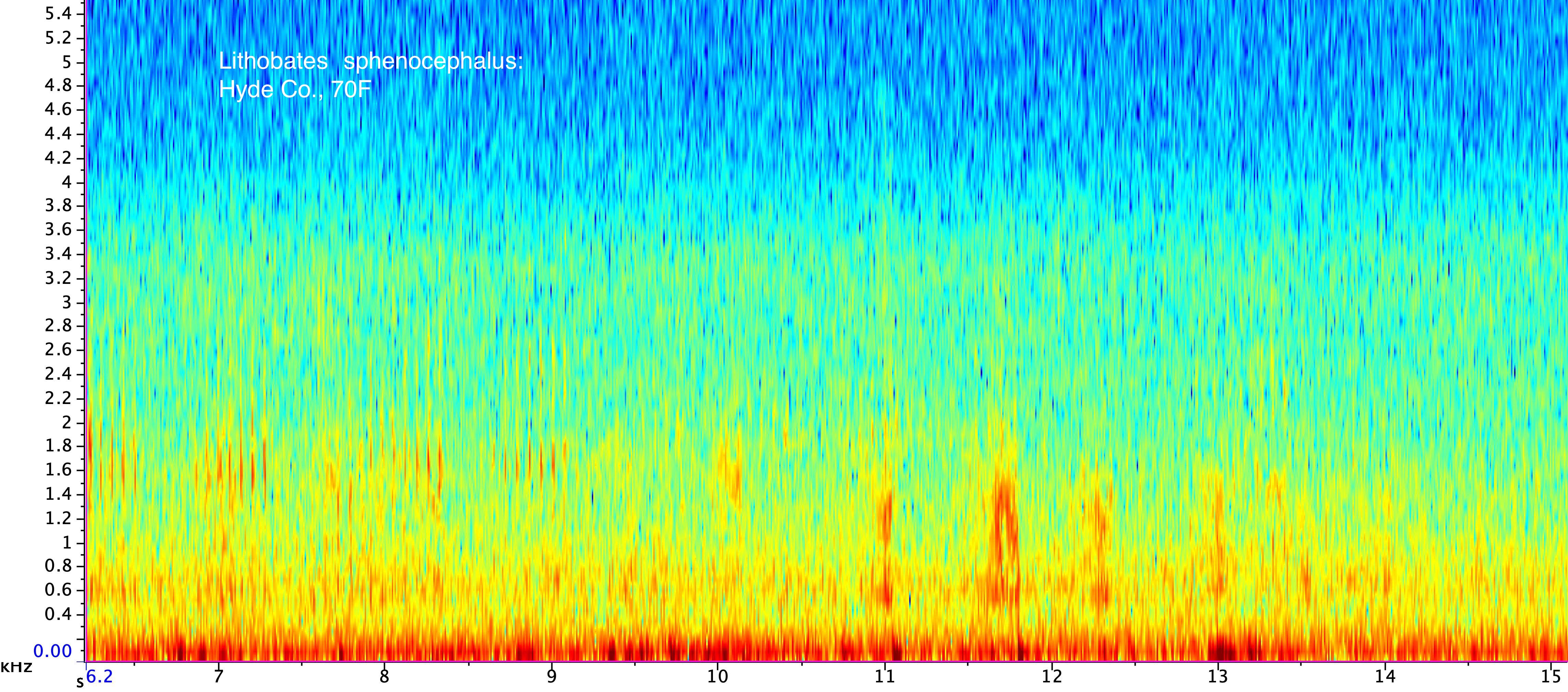
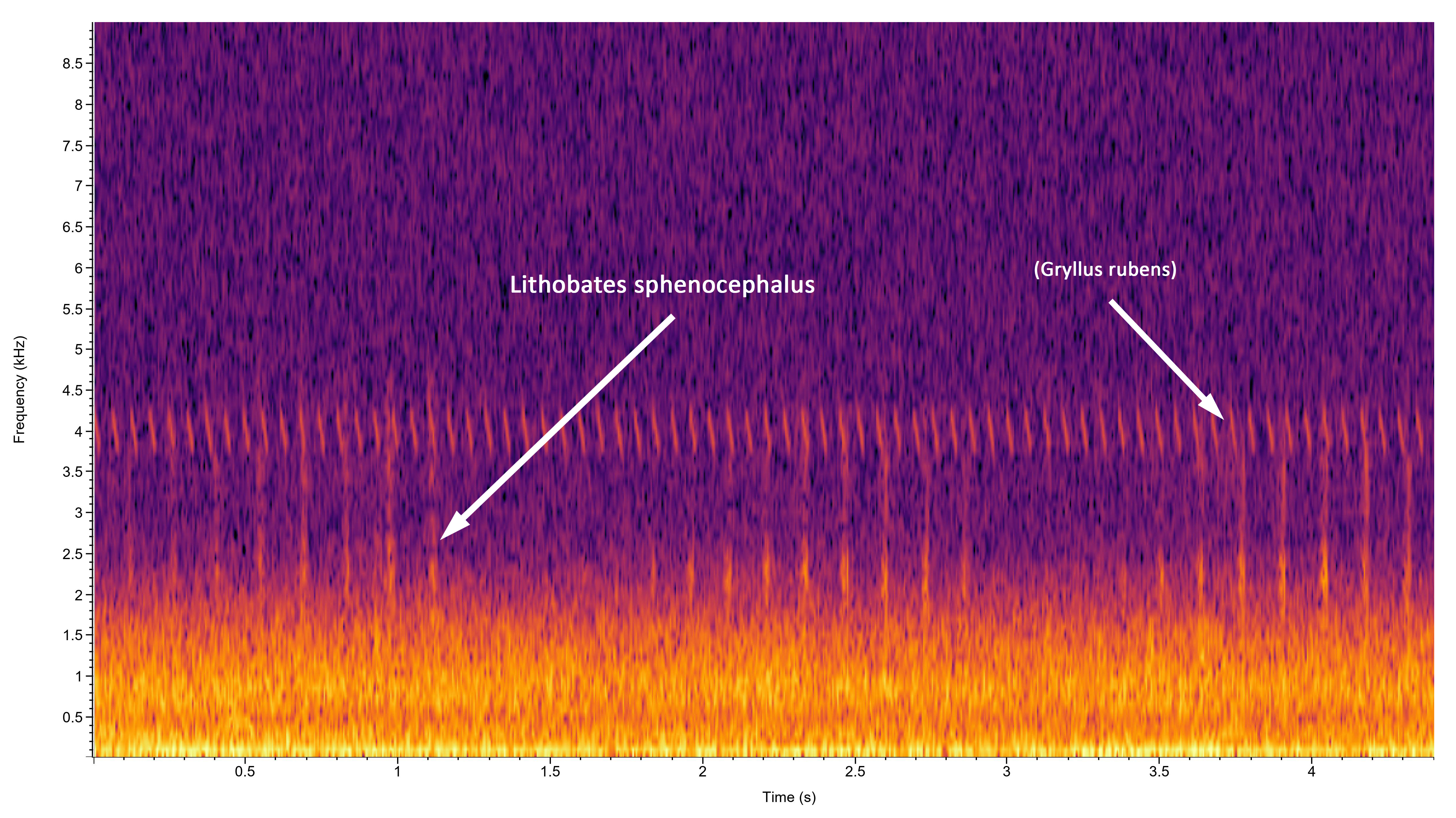
Description: The adults are medium-sized frogs with a dorsal ground color that can be various shades of dark olive-green, brown, bronze, gray or light green. Some specimens may be bicolored, with green between the dorsolateral folds and brown along the sides of the body (Dodd 2013). A conspicuous yellowish-white dorsolateral fold in present that extends from behind each eye to near the attachment of the rear legs, and a white to dull yellowish spot is present in the center of the tympanum. The skin on the dorsum is mostly smooth except for scattered small tubercles and a series of short, irregular raised ridges between the dorsolateral folds. The snout is rather pointed relative to many of our native ranids, and there is a light line on each side of the upper jaw that projects rearward towards the insertion of the front leg. Dark mottling is usually evident below the light line and on the opposing top of the lower jaw. There is a series of large brown or blackish spots on the back, along the sides, and on the legs. These can be rounded, oval, or more elongated, and are sometimes arranged in two or three irregular rows between the dorsolateral folds. The spots along the sides tend to become progressively smaller towards the belly. The hind legs are long with webbed toes, and the belly, chest and undersides of the legs are white. The hidden portion of the back of the hind leg is variable, but in most specimens it is predominantly light with large, connected blotches of dark pigment. L. sphenocephalus and L. kauffeldi overlap in the northern Coastal Plain of North Carolina. Details concerning how to distinguish between these species are in the account of L. kauffeldi . Lithobates kauffeldi can be difficult to distinguish from L. sphenocephalus and is most reliably separated using the advertisement calls (see the detailed discussion in the L. kauffeldi account). These species only occur sympatrically in the northeastern corner of the state so that most specimens can be reliably assigned to species based on the collection site. Vocalizations: The advertisement call is distinctive, but rather complex and difficult to describe. It generally consists of one or more rapid sequences of 2-6 guttural notes or clucks ('ah-ah-ah-ah' or 'aka-aka-aka-aka') that is often referred to as a chuckle call sequence. These may be preceded or followed with a series of longer guttural groans that some have likened to running a thumb over an inflated balloon. Numerous variations on this general pattern occur. Individuals also may issue a release call (short trill) if a male is amplexed by another male, and a distress call when ceased by a predator such as a snake that sounds like a guttural scream. This may function to draw other predators to the attacker. Technical Reference: Dodd (2013)Online Photos: Google iNaturalist Observation Methods: Individuals can be found calling at the breeding sites and moving across roads on rainy nights. Searching areas in or near the shorelines of ponds or other wetlands can be productive.
Download Video:
"MP4"
Download Video:
"MP4"
AmphibiaWeb Account
Distribution in North Carolina
Distribution Comments: The Southern Leopard Frog has a large range that extends from much of New Jersey and the Delmarva Peninsula southward through the Atlantic Coastal Plain and Piedmont of the southeastern United States to southern Florida. From there the range extends westward through all of Georgia, Alabama, Mississippi, Louisiana, and Arkansas to eastern Texas and eastern Oklahoma. The range extend northward from the Gulf States to encompass Tennessee, most of Kentucky, extreme southern Ohio, southwestern Indiana, southern Illinois, most of Missouri, and southeastern Kansas. Schlesinger et al. (2018) noted that previous records from southern New York and northern New Jersey are not valid, while Beane et al. (2010) noted that this species extends into the mountains of South Carolina. In North Carolina, populations occur throughout the Coastal Plain and eastern and central Piedmont. They are absence from the mountains and most of the western Piedmont.Distribution Reference: Beane et al. (2010), Dodd (2013), Schlesinger et al. (2018)County Map: Clicking on a county returns the records for the species in that county.
GBIF Global Distribution
Key Habitat Requirements
Habitat: The juveniles and adults are typically found in mesic settings, and often in landscapes that have clusters of wetlands of various hydroperiods that are embedded within forests (Dodd 2013). They can use both upland and bottomland habitats, and both deciduous and pine forests, as long as there are moist and humid microhabitats available. Individuals are often found in close proximity to the breeding sites or other bodies of water, but may forage in the surrounding habitats. Breeding occurs in both seasonal and permanent bodies of water. Dodd (2013) listed a variety of known multipurpose habitats that are used for breeding, terrestrial habitats, and overwintering sites. These include riparian floodplains along rivers and streams, wooded ravines, cypress savannas, cypress-gum ponds, dome swamps, Carolina bays, Atlantic White Cedar swamps, mangrove forests, sloughs, wet meadows, marshes, lake margins, beaver ponds, and artificial habitats such as canals and ditches. Individuals have also been found in freshwater tidal wetlands and sawgrass marshes. Environmental and Physiological Tolerances: The eggs and larvae appear to be moderately salt tolerant. The larvae have been found in coastal, salt-invaded wetlands in North Carolina at concentrations as high a 11 ppt (Albecker and McCoy 2017). Adults have been found in a variety of other salt-invaded wetlands such as coastal marshes, creeks, and ponds with saline contents of 0.05 to 21.4 ppt (Dodd 2013), although salinities as high as 21.4 ppt would undoubtedly be lethal to the eggs and larvae (Hopkins and Brodie 2015). Christman (1974) found that adults from coastal populations are more salt-tolerant than those from more inland localities. Biotic Relationships: The eggs and larvae are commonly found in seasonal ponds with moderate to long hydroperiods and are vulnerable to predation from both invertebrate and vertebrates. Richter (2000) found that the eggs and developing embryos at a site in southern Mississippi were heavily preyed upon by phryganid caddisflies. Johnson et al. (2003) discovered that late-term embryos will hatch earlier when in the presence of crayfish (Procambarus nigrocinctus ) and dytiscid beetle larvae that feed on the developing embryos. Responses were strongest to the crayfish, which was a far more effective predator on the embryos. The Eastern Newt is also a well documented predator on the eggs of this and other amphibian species, and American Bullfrog tadpoles can kill and eat the hatchlings (Boone et al. 2004). Notophthalmus perstriatus and N. viridescens ), Ambystoma larvae (Stemp et al. 2021), fishes (particularly centrarchids), aquatic and semiaquatic snakes (Agkistrodon ; Nerodia ; Regina ; Thamnophis ), snapping turtles (Chelydra ), wading birds, grackles, raccoons, and foxes. Fogarty and Hetrick (1973) recovered 25 adults from the guts of Cattle Egrets in Florida.
Life History and Autecology
Breeding and Courtship: As might be expected for this wide-ranging species, the seasonal patterns of calling and breeding vary geographically. Peak breeding for southern populations in Louisiana, Mississippi, Alabama, Florida and elsewhere in the Deep South typically occurs during the cooler months of late fall through late winter or early spring. However, sporadic breeding can occur at other times of the year, particularly following heavy rain events (Caldwell 1986, Dodd 2013, Krysko et al. 2019, McCallum et al. 2004, Mount 1975, Dundee and Rossman 1989). In Georgia, breeding mostly occurs from December-February in the southern part of the state, and from March-June in northern Georgia (Jensen et al. 2008). Breeding at a site in Mississippi that was intensely studied by Doody and Young (1995) was episodic, with widely-spaced breeding bouts occurring from November through March after heavy rains. The beginning of fall and early winter breeding for some local populations may be constrained by when seasonal ponds first fill with the onset of cooler weather and decreased evapotranspiration rates.Reproductive Mode: Females deposit their eggs in large globular masses. Each female normally lays a single egg mass that is either attached to a support structure or simply laid freely on mats of submerged plant debris or other substrates. Dodd (2013) noted a general tendency for the masses to be laid in shallow, sunny microhabitats when water temperatures are relatively cool (e.g., winter), and in deeper waters when conditions are warmer. Females may either scatter their egg masses wide apart in a pond, or lay them communally in groups (Caldwell 1986). Trauth (1989) found a group of 75 masses in close contact to one another in Arkansas, while Caldwell (1986) found clusters of 6-126 masses in South Carolina. Communal deposition such as this tends to prevail during the cooler months of the year. Its exact function is unknown for this species, but Caldwell (1986) surmised that clusters of dark egg masses may function as thermal traps that allow more rapid embryonic development. Aquatic Life History: In situations where the eggs are laid communally, the hatchlings often congregate in close proximity for several days before dispersing widely throughout the breeding sites. The larval diet is poorly documented but likely includes both plant and animal material, including algae, zooplankton, and possibly small insects and other invertebrates. Hillis (1982) found that the larvae mostly filter-fed on green algae at a site in Texas, while Watters et al. (2018) found that the tadpoles were effective predators on mosquito larvae. Terrestrial Life History: Both the young metamorphs and adults often move substantial distances from the breeding sites as they make their way to terrestrial habitats. Movements typically occur during wet weather and individuals seek out moist, humid environments for permanent settlement. Others may remain near aquatic sites, particularly where there is thick shoreline vegetation that provides good cover and opportunities for foraging (Kilby 1945). Dodd (2013) noted that forests with well-developed herbaceous understories and low overhead canopies often provide ideal habitats. Desmognathus salamanders and small frogs, including Acris gryllus , Anaxyrus quercicus , Hyla cinerea , H. squirella , and Pseudacris ocularis (Crawford et al. 2009, Dodd 2013, Duellman and Schwartz 1958, Jensen et al. 2008, Kilby 1945). Some of the invertebrate prey that are consumed (summarized by Dodd 2013) include roaches, beetles, orthopterans, true bugs, ants, flies, mayflies, lepidopteran larvae, grasshoppers, spiders, snails and crayfishes.
General Ecology
Population Ecology: Very limited information is available on population structure and population interconnectivity across the landscape. The adults are known to move long distances from the breeding sites and often quickly colonize created wetlands. This suggests that a single local population may use clusters of local ponds with high connectivity among ponds. McKee et al. (2017) analyzed genetic variation in local pond sites in an 11,800-ha site in Georgia and found little evidence of significant population structure at this spatial scale. Local populations did exhibit genetic isolation by distance, but the pattern was weaker when compared to that of Eurycea quadridigitata that was sampled at the same site.Community Ecology: Larvae of the Southern Leopard Frog often share breeding ponds with numerous other amphibians, including both potential predators and competitors. Attempts to understand ecological interactions between these species have relied almost entirely on studies in laboratory aquaria or outdoor tanks with experimentally assembled communities. Studies by Morin (1983) using artificial pools demonstrate that interactions between community members are often mediated by predators, particularly the Eastern Newt. In the absence of predators, L. sphenocephalus competed relatively well with other anuran species that shared tanks. Newts played an important role in reducing the densities of competitors, mediating density-dependent competition, and influencing whether L. sphenocephalus tadpoles overwintered in the experimental tanks.L. sphenocephalus can depress the growth rates of other anurans, including Hyla gratiosa , H. chrysoscelis , and Hyla andersonii (Alford 1989a, Morin 1983, Pehek 1995). The relative time at which anurans are added to pools can also influence the outcome of competitive interactions (Alford 1989a, Alford and Wilbur 1985). We have much to learn about competitive and predatory interactions in natural breeding sites, and the extent to which results from studies of artificial communities apply to natural systems.
Adverse Environmental Impacts
Habitat Loss: The Southern Leopard Frog has undoubtedly suffered significant historical losses since European colonization due to the loss of natural wetlands and widespread deforestation of many areas of the Southeast. Habitat Fragmentation: Southern Leopard Frog appears to tolerate habitat fragmentation fairly well so long as suitable breeding sites are scattered across the landscape and habitats are available for the juveniles and adults (Dodd 2013). This species is generally poorly represented in highly urbanized settings.
Status in North Carolina
NHP State Rank: S5Global Rank: S5Environmental Threats: Populations in North Carolina are threatened locally by urbanization, vehicular losses, and the conversion of forested habitats to agricultural fields and other uses.Status Comments: The southern Leopard Frog tolerates landscape disturbances relatively well and its use of artificial aquatic habitats has allowed it to adapted to human-altered landscapes. Although this species has been adversely impacted by the historical loss of natural wetlands and adjoining forested uplands, populations in North Carolina appear to be stable and show no evidence of recent widespread population declines. Stewardship: Populations are best maintained by have a cluster of seasonal or semi-permanent breeding sites that are surrounded by forests with mesic microhabitats.

 »
»