Amphibians of North Carolina
Home Page
Recent Entries
Recent Account Updates
General Search
Submit a Public Record
References
Maps
Checklist
NC Biodiversity Project
Comments
Scientific Name:
Common Name:
Family (Alpha):
AMBYSTOMATIDAE
AMPHIUMIDAE
BUFONIDAE
CRYPTOBRANCHIDAE
HYLIDAE
MICROHYLIDAE
PLETHODONTIDAE
PROTEIDAE
RANIDAE
SALAMANDRIDAE
SCAPHIOPODIDAE
SIRENIDAE
«
Home
»
Plethodontidae Members:
Aneides aeneus
Aneides caryaensis
Desmognathus adatsihi
Desmognathus aeneus
Desmognathus amphileucus
Desmognathus anicetus
Desmognathus aureatus
Desmognathus bairdi
Desmognathus balsameus
Desmognathus campi
Desmognathus carolinensis
Desmognathus conanti
Desmognathus folkertsi
Desmognathus fuscus
Desmognathus gvnigeusgwotli
Desmognathus imitator
Desmognathus intermedius
Desmognathus kanawha
Desmognathus lycos
Desmognathus marmoratus
Desmognathus mavrokoilius
Desmognathus monticola
Desmognathus ocoee
Desmognathus orestes
Desmognathus organi
Desmognathus perlapsus
Desmognathus planiceps
Desmognathus santeetlah
Desmognathus tilleyi
Desmognathus unidentified species
Desmognathus valtos
Desmognathus wrighti
Eurycea arenicola
Eurycea chamberlaini
Eurycea cirrigera
Eurycea guttolineata
Eurycea junaluska
Eurycea longicauda
Eurycea quadridigitata
Eurycea unidentified species
Eurycea wilderae
Gyrinophilus porphyriticus
Hemidactylium scutatum
Plethodon amplus
Plethodon aureolus
Plethodon chattahoochee
Plethodon cheoah
Plethodon chlorobryonis
Plethodon cinereus
Plethodon cylindraceus
Plethodon glutinosus
Plethodon hybrids
Plethodon jacksoni
Plethodon jordani
Plethodon meridianus
Plethodon metcalfi
Plethodon montanus
Plethodon richmondi
Plethodon serratus
Plethodon shermani
Plethodon teyahalee
Plethodon unidentified species
Plethodon ventralis
Plethodon welleri
Plethodon yonahlossee
Plethodon yonahlossee population 1
Pseudotriton montanus
Pseudotriton ruber
Pseudotriton ruber nitidus
Pseudotriton ruber ruber
Pseudotriton ruber schencki
Stereochilus marginatus
Desmognathus perlapsus
- Chattooga Dusky Salamander
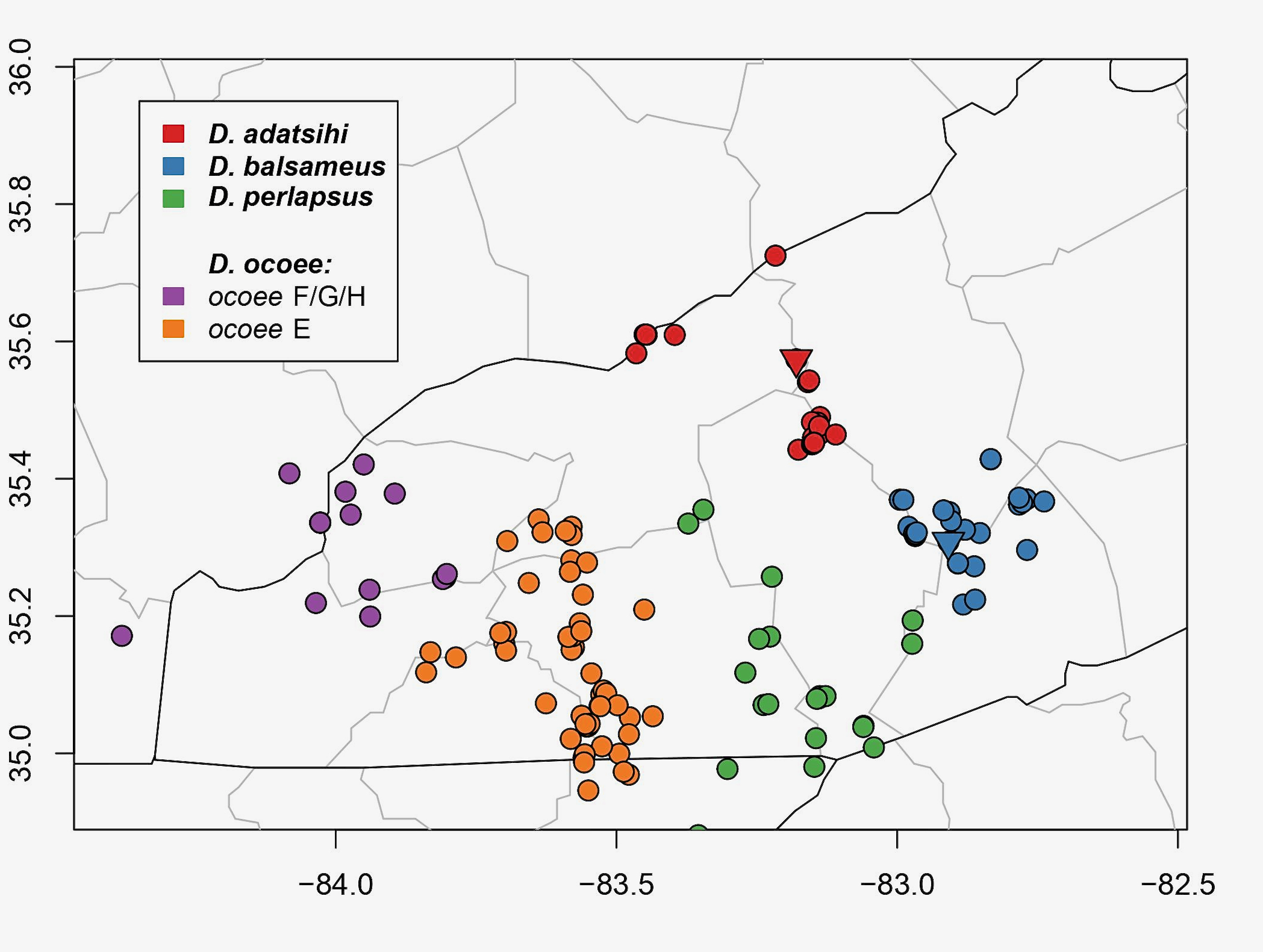
caption
A map of the D. ocoee complex based on genetically identified specimens (courtesy of Alex Pyron).
Taxonomy
Class:
Amphibia
Order:
Caudata
Family:
Plethodontidae
Subfamily:
Plethodontinae
Taxonomic Comments:
Members of the genus
Desmognathus
are commonly known as dusky salamanders because of their overall dark brown or dusky ground color. Like many plethodontid salamanders, they have proven to be a taxonomically challenging group that contains several species complexes. Kozak et al. (2005) documented 35 major lineages in the eastern US, even though only 22 species were formally recognized by taxonomists in 2021. This suggests that there are numerous cryptic species that remain to be described. A more recent comprehensive molecular survey of populations in the eastern US by Beamer and Lamb (2020) indicate that at least 45 major lineages or clades are present.
In North Carolina, a group of medium-sized dusky salamanders with rounded, unkeeled tails (
D. carolinensis
,
D. imitator
,
D. ocoee
, and
D. orestes
) have been treated as members of an informal cryptic species complex known as the
D. ochrophaeus
complex (Tilley and Mahoney 1996). The members of this group are commonly referred to as mountain dusky salamanders, and were previously treated as a single species (
D. ochrophaeus
, sensu lato) before being split. The most recent molecular study based on mtDNA sequence data (Beamer and Lamb 2020), as well as previous studies (see Beamer and Lamb 2020), indicate that there are many additional clades within the
D. ochrophaeus
complex that may represent undescribed cryptic species. In addition, certain forms appear to be more genetically similar to
D. fuscus
than to other members of the
D. ochrophaeus
complex. Some of the highlights of this study are that 1) Populations currently referred to as
D. carolinensis
do not appear to constitute a genetically distinct lineage and instead cluster with 39 populations of
D. fuscus
, 2) Populations currently referred to as
D. ocoee
contain several clades that may reflect additional cryptic species; in some cases these clades contain mixtures of populations that are currently referred to as
D. ocoee
,
D. conanti
and
D. apalachicolae
, 3) Populations currently referred to as
D. orestes
contain three clades, one of which is sister to a clade containing
D. ochrophaeus
and other clades of
D. orestes
, and 4) Populations currently referred to as
D. imitator
constitute a well-defined clade that is not closely related to other members of the
D. ochrophaeus
complex and should not be treated as a member of that complex. One problematic issue with studies of this sort is that the results based on using different molecular evidence (e.g., mtDNA; nuclear DNA; allelic frequencies) are sometimes discordant, in part due to past or present hybridization that commonly occurs between closely related plethodontid species (see Pyron et al. 2020) as it relates to
Desmognathus
species).
Pyron and Beamer (2022a) conducted additional molecular and morphometric studies and recognized five species within the
oconee
group. These include two new species (
D. adatsihi
;
D. balsameus
) and one resurrected species (
D. perlapsus
). Four of the five species in the
D. oconee
complex occur in western North Carolina and are best identified using their geographic ranges (see map above).
Species Comments:
Identification
Description:
Desmognathus perlapsus
is a medium-size
Desmognathus
and closely resembles other members of the
D. ocoee
complex. It is best identified by range and the collection site. The tail is mostly rounded but tends to become more flattened on the distal portion. The dorsum typically has 4-6 pairs of dorsal spots between the limbs that become fused with age to produce a wavy pattern with dark margins around the spots. White flecking is often present on the upper surfaces, and whitish mottling on the sides and venter. Some populations in the Cowee Mountains and Highlands Plateau also occasionally exhibit red, orange, or yellow cheek patches (Pyron and Beamer, 2022a). This species lacks corified, dark toe tips as seen in most of the more aquatic
Desmognathus
species.
Males in local populations of this and other members of the
ocoee
complex typically average slightly larger than females and are more likely to be melanistic. Relative to females, the males have well developed premaxillary teeth and large jaw musculature. They also possess small mental glands and have a mouth line that appears sinuate when viewed from the side. The hatchlings of the coee complex vary from about 13-18 mm TL and have prominent yolk reserves. The larvae have rounded snouts, very small whitish gills, and a series of 4-6 pairs of alternating light spots along the dorsum.
Online Photos:
Google
iNaturalist
Observation Methods:
Individuals are easy to collect by searching beneath cover objects along stream margins, under rocks and debris in the stream proper, or in the adjoining woods. They are most easily observed at night when they actively forage and seek mates on the forest floor.
AmphibiaWeb Account
Distribution in North Carolina
Distribution Comments:
Desmognathus perlapsus
ranges from the Alarka and Cowee Mountains in western North Carolina southward through the western headwater mountain streams of the Savannah River drainage in northwestern South Carolina, northeastern Georgia, and adjacent North Carolina. From there the range extends southwestward into the Chattahoochee River drainage in the Piedmont of Georgia and Alabama to the Fall Line and adjacent Coastal Plain in southeastern Alabama (Pyron and Beamer, 2022a). In North Carolina, this species occurs between the Tuckasegee and Little Tennessee Rivers in extreme eastern Clay County, extreme western Macon County, extreme southwestern Transylvania County and extreme southern Swain County.
County Map:
Clicking on a county returns the records for the species in that county.
GBIF
Global Distribution
Key Habitat Requirements
Habitat:
In western North Carolina.
D. perlapsus
is associated with montane hardwood forests with cool, clear-flowing headwater streams and seepages and wet rock faces. Brooding females and overwintering juveniles and adults congregate in aquatic habitats, but non-brooding individuals move away from aquatic habitats and live on the forest floor during the warmer months of the year. At sites where mesic conditions prevail, they can be found well away from streams and other aquatic sites. Farther south, populations in Georgia and Alabama are most commonly found in the immediate vicinity of streams due to less mesic conditions.
Biotic Relationships:
This and other members of the
D. ocoee
complex are palatable to predators and are undoubtedly preyed upon by aquatic predators such as crayfishes, as well as woodland birds, small snakes, large salamanders, and small mammals such as shrews that live in stream, streamside, and forest-floor habitats in mesic forests. The Spring Salamander (
Gyrinophilus porphyriticus
), members of the Black-bellied Salamander complex, and the Seal Salamander (
D. monticola
) feed on members of the
D. ochrophaeus
and
D. ocoee
species complex (Bruce 1979, Coker 1931, Formanowicz and Brodie 1993, Hairston 1986, Petranka 1998, Whiteman and Wissinger 1991) and all three undoubtedly feed on
D. perlapsus
as well.
Life History and Autecology
Breeding and Courtship:
The courtship behavior of members of the
D. ocoee
complex appear to be identical. The breeding season is also similar other than constraints due to local climatic conditions associated with different elevations in the Blue Ridge Mountains. The adults mate during the warmer months of the year in spring, late-summer, and autumn, and the females reproduce annually in most populations (Forester 1977, Huheey and Brandon 1973, Martof and Rose 1963). Courtship involves a stereotypic tail straddle walk, but detailed descriptions of courtship behavior are not available. The basic sequence of events involves a period of persuasion that is followed by a tail straddle walk and spermatophore deposition. The male may repeatedly snap or pull his chin across the female's dorsum, which scratches the female's skin and transfers mental gland secretions into the female's circulatory system (Houck 1986). Mental gland secretions act to enhance the sexual responsiveness of females to males (Houck and Reagan 1990).
In this and other members of the
D. ocoee
complex, females often court several times per year and are capable of long-term sperm storage. As such, sperm competition likely occurs among males (Petranka 1998). When two males are near a female, the larger will often chase the smaller one away prior to mating. Each male normally deposits only one spermatophore per night, even when experimentally presented with multiple partners (Verrell 1988a). In addition, females are not responsive to males for 2-4 days after successful insemination. This corresponds to a time when the female's cloaca is physically blocked by the sperm cap (Verrell 1988b, 1991).
Reproductive Mode:
The nesting biology of members of the
D. ocoee
complex -- as well as
D. carolinensis
and
D. orestes
-- appear to be very similar or identical. Some of the studies that reported on the nesting biology prior to the splitting of
D. ochrophaeus
(sensu lato) and
D. ocoee
(sensu lato) into multiple species (see above) are confounded because they involved two or more species (e.g., Martof and Rose, 1963). The following provides a general summary of the nesting biology of this group of closely related species, with references to specific species, including
D. perlapsus
, where possible.
Females often move from streamside or forested habitats to their nesting sites 1-3 weeks before ovipositing. Each female lays a small grape-like cluster of eggs. These are typically placed in a hollowed depression that is constructed beneath decaying logs, rocks, leaf litter or moss mats in or near springs, seeps, and small streams (Petranka 1998). Each egg has a short gelatinous stalk and the stalks become twisted around each other to form a single cluster that the female broods. Mature ova average about 2-3 mm in diameter. The mean clutch size of 21 populations with mixed species that were studied by Martof and Rose (1963) varied from 8-20 eggs, while nine clutches of
D. perlapsus
collected by Bruce (1990) in western North Carolina contained from 8-23 eggs and averaged 14. The modal clutch sizes for two populations of
D. ocoee
(sensu stricto) studied by Bernardo (1994) were 12 and 16 eggs, while Tilley (1972) reported clutches of 10-19 eggs (mean = 14 eggs) for
D. perlapsus
and 13-24 (mean = 18) for
D. carolinensis
on Mt. Mitchell. Clutch size is positively correlated with female body size in this and most other
Desmognathus
species and increases with elevation (Petranka 1998). Forester (1977) collected over 400 females of
D. perlapsus
from mid-June through late-August that were brooding their eggs beneath decaying logs, rocks, leaf litter and moss mats in a small stream in Macon County. Many (probably most) of the females reproduced annually and they returned to the same stream stretch to breed.
Females of all members of the Mountain Dusky Salamander complexes remain with their clutches through hatching. Brooding functions to protect the embryos from predators such as carabid beetles and other salamanders. It also functions to remove dead eggs from nests, to reduce egg desiccation, and to reduce fungal attacks (Petranka 1998, Tilley 1972). Females of
D. perlapsus
(and presumably other members of the
D. ocoee
complex) will lunge at and bite certain intruders, and their aggression towards intruders increases during the brooding season as their investment in parental care increases (Forester 1983). Clutches at or near hatching have been found from August to late September. The estimated incubation periods for North Carolina populations of
D. perlapsus
were 57-74 days (Forester 1977) and 50-60 days (Tilley 1972), and 52-69 days for
D. carolinensis
on Mt. Mitchell (Tilley 1972). Embryonic survivorship from early developmental stages to hatching is near 80% in clutches examined by Tilley (1972).
Females of members of the Mountain Dusky Salamander complexes greatly reduce their food intake when brooding and appear to feed opportunistically on prey that approach the brooding sites (Forrester 1977, Martof and Rose 1963, Tilley 1972). Most females have a negative energy budget and lose weight over the brooding season. Brooding females sometimes feed on their own eggs, but the extent to which this reflects the attack of healthy embryos versus the removal of dead or diseased eggs from nests is not fully understood. Tilley (1972) found that females will preferentially eat dead eggs introduced into their clutches, but would sometimes eat healthy eggs from their own broods immediately after consuming a dead egg. Brooding females that are disturbed will consume healthy eggs from their own clutches, and hatchlings are sometimes cannibalized (Petranka 1998). Collectively, these data suggest that females regularly remove dead eggs from their clutches, but occasionally eat healthy eggs and hatchlings. Perhaps the movement of hatchlings or late-term embryos within the egg capsule stimulates hungry females to prey upon their offspring.
Aquatic Life History:
Fully developed embryos of members of the
D. ocoee
complex typically hatch in North Carolina in August through September. The larvae subsist on small invertebrates and the larval period last less than one year. Bruce (1989) reported that the larvae of
D. perlapsus
metamorphose in May and June of the following year when 11-15 mm SVL. Beachy (1995) grew larvae of
D. ocoee
in the laboratory at temperatures that resembled those in natural streams and found that the larval period typically lasted around 9-10 months. His experimental manipulation of food levels and water temperatures indicate that both can affect larval growth and development rates.
Terrestrial Life History:
Members of the
D. ocoee
and
D. ochrophaeus
complexes have very similar terrestrial life histories. Although the terrestrial ecology of
D. perlapsus
is poorly documented, it presumably is very similar to that of other members of the mountain dusky salamander complexes. A general summary of the terrestrial life history of these species follows.
After metamorphosing the young juveniles disperse from the nesting sites and move into either streamside habitats or the surrounding forest floor where they feed during the warmer months of the year. The juveniles and adults are generalist feeders that consume a wide variety of invertebrates, including many types of larval and adult insects, spiders and mites, nematodes, salamander eggs and juveniles, and shed skin (Petranka 1998). Individuals are active on the ground surface except during the coldest winter months when the adults and juveniles congregate in seepages or underground retreats.
The juveniles and adults are nocturnally active. They hide beneath cover objects or in recesses during the day, then emerge at night to feed or find mates. In the southern Appalachians individuals frequently climb plants or tree trunks on rainy nights, but are rarely encountered more than a meter or so above ground level. The adults are territorial and will actively defend space from conspecifics, particularly intruding males (Petranka 1998). Large males tend to dominate smaller males, and may bite them if the opponent does not flee when approached. Bites are often directed to the tail, and on rare instances, a portion of the tail may be lost. Population densities on rockfaces usually exceed those of streamside habitats, perhaps because of reduced predation from larger salamanders that co-occur in stream and streamside communities.
Growth rates vary depending on age, elevation, and the microhabitats frequented by the juveniles and adults. Males of two rockface populations of
D. perlapsus
that were studied by Tilley (1977, 1980) grew about 5-7 mm per year SVL until age 4-5, then slowly declined thereafter. The pattern was similar for females, except that their growth rates begin to decline after reaching 3-4 years of age. Individuals in western North Carolina and extreme southeastern Tennessee often become sexually mature when < 30 mm SVL (Petranka 1998). Bruce (1990) found that both sexes of
D. perlapsus
reach sexual maturity in a stream population when 3-years old, and that females first oviposit when 4-years old. In general, the age and size at first reproduction, maximum adult size, mean clutch size, and adult survivorship tend to increase with elevation (Tilley 1977, 1980).
General Ecology
Adverse Environmental Impacts
Status in North Carolina
NHP State Rank:
[S4]
Global Rank:
GNR
Status Comments:
Desmognathus perlapsus
ranges from the Alarka and Cowee Mountains in western North Carolina southward through northeastern Georgia into the Piedmont of Georgia and Alabama, and to the Fall Line and adjacent Coastal Plain of southeastern Alabama. In North Carolina, this species occurs between the Tuckasegee and Little Tennessee Rivers in extreme eastern Clay County, extreme western Macon County, extreme southwestern Transylvania County, and extreme southern Swain County. Local populations are found in stream and streamside habitats in headwater and lower-order streams. Many local populations in North Carolina are in national forests and other lands that offer a significant degree of protection.
Photo Gallery for
Desmognathus perlapsus
- Chattooga Dusky Salamander
5 photos are shown.
Recorded by: David George
Swain Co.
Recorded by: David George
Swain Co.
Recorded by: A. Coburn
Transylvania Co.
Recorded by: A. Coburn
Transylvania Co.
Recorded by: Jim Petranka
None Co.
Comment: A map of the D. ocoee complex based on genetically identified specimens (courtesy of Alex Pyron).

 »
»