Amphibians of North Carolina
Home Page
Recent Entries
Recent Account Updates
General Search
Submit a Public Record
References
Maps
Checklist
NC Biodiversity Project
Comments
Scientific Name:
Common Name:
Family (Alpha):
AMBYSTOMATIDAE
AMPHIUMIDAE
BUFONIDAE
CRYPTOBRANCHIDAE
HYLIDAE
MICROHYLIDAE
PLETHODONTIDAE
PROTEIDAE
RANIDAE
SALAMANDRIDAE
SCAPHIOPODIDAE
SIRENIDAE
«
Home
»
Hylidae Members:
Acris crepitans
Acris gryllus
Hyla andersonii
Hyla chrysoscelis
Hyla cinerea
Hyla femoralis
Hyla gratiosa
Hyla squirella
Hyla versicolor
Pseudacris brimleyi
Pseudacris collinsorum
Pseudacris crucifer
Pseudacris feriarum
Pseudacris nigrita
Pseudacris ocularis
Pseudacris ornata
NC
Records
Hyla femoralis
- Pine Woods Treefrog
caption
A view of the rear thighs with bright yellow-orange spotting.
Taxonomy
Class:
Amphibia
Order:
Anura
Family:
Hylidae
Subfamily:
Hylinae
Taxonomic Comments:
Duellman et al. (2016) elected to split treefrogs in the genus
Hyla
into two genera.
Hyla
(sensu stricto) refers to a group of species that are found in Eurasia, while a new genus
Dryophytes
was resurrected to include all of the North and Central American species and three other species that are found in eastern temperate Asia. Members of these proposed genera cannot be distinguished by any known morphological features, but represent two clades. Whether members of these clades should be placed in separate genera or treated as lesser taxa such as subgenera is arbitrary and dependent on one's taxonomic philosophy. Here, we retain
Hyla
for North American species to be consistent with current usage by the Society for the Study of Amphibians and Reptiles.
Species Comments:
Identification
Description:
The Pine Woods Treefrog is a small frog with moderately developed toe pads and rear thighs with bright yellow-orange spots on a dark background. Most adults have a dark blotch between the eyes, and many have a gradually widening dark line on each side that begins at the nostril and extends through the eye and tympanum. It continues onto the lower side where it broadens to become a diffuse band, with the dorsal margin often delineated by a thin dark line. The dorsal ground color is usually grayish, tan, brown or reddish brown, but individuals are commonly seen that are light green, greenish gray or dark brown. The ground is usually overlain with a series of darker patches or blotches on the back and upper legs. These may be obscure in some individuals, or be well developed and even anastomose to varying degrees in others. Individuals can change color and tend to become lighter at night. The venter is white and unspotted except for a dark vocal sac on sexually active males. The young metamorphs are often green and lack the distinctive marks of the adults. They can be difficult to distinguish from the young of other hylids such as
Hyla squirella
and
H. cinerea
(Wright 1932).
The males typically average around 8-12% smaller than the females based on SUL. Some of the values that were reported by Dodd (2013) are a mean of 27.1 mm SUL for males and 31.1 mm SUL for females from central Florida, and 30.7 mm SUL for males and 34.2 mm SUL for females from South Florida. Reported size ranges for the adults are 19–40 mm SUL (mean 32 mm) for specimens from northern Florida and 28–41 mm SUL (mean 33 mm) for Alabama specimens. Beane et al. (2010) list a range of 25-38 mm for specimens in Virginia and the Carolinas versus 25-45 mm for Georgia specimens (Jensen et al. 2008).
The very small larvae are brownish yellow with a distinct lateral stripe and sharply bicolored tail musculature (Dodd 2013). As they age the dorsolateral body stripe is lost, the tail stripe becomes more distinctive, and the body assumes a dark olive to black coloration with a light (often bright yellow) venter. A pale postorbital stripe is present, but there is no interorbital bar (Dodd 2013). The tail musculature on older larvae has a distinct stripe, the tail fin is flecked or blotched towards the margins, and there is a well-developed flagellum at the tip of the tail. The full grown tadpoles can reach about 22–38 mm TL (Brown 1956, Jensen et al. 2008). Much like
Hyla chrysoscelis
, this species has inducible defenses. When exposed to certain predators the older larvae develop relatively short tails with conspicuous orange-red coloration and blackish blotching. So, expect substantial variation depending of local site conditions.
Vocalizations:
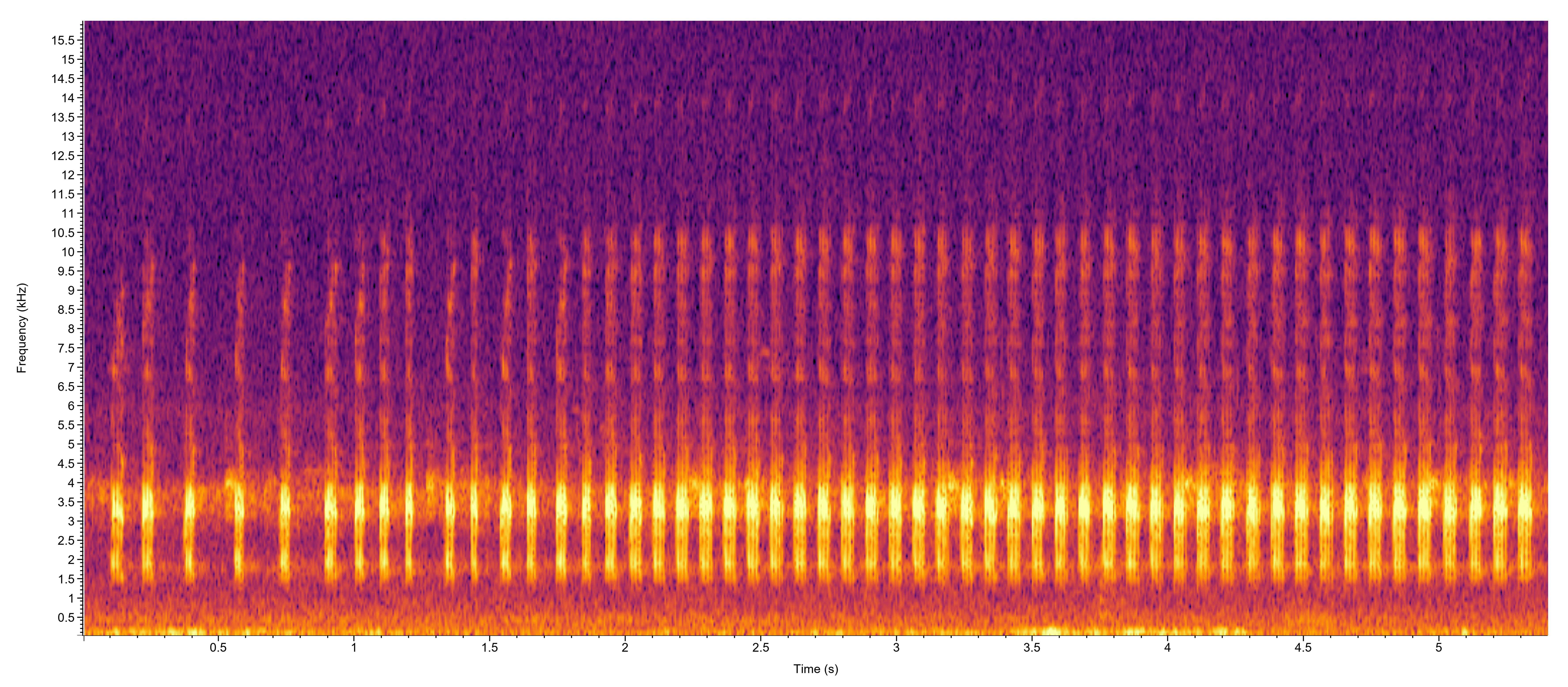
Dodd (2013) noted that the males have two distinctive calls. One is made from the trees and occurs throughout the warm activity season, while the other is a more typical male advertisement call that is made at the breeding sites. The tree call is similar to the advertisement call, but differs in the number of pulses per call, the time between pulses, the pulse repetition rate and the minimum call frequency (Schad 2001). The tree call requires less energy than the advertisement call and can occur both during the day and night. However, it is most commonly issued at dawn. Surprisingly, Schad (2001) found that the tree call is not given more often under environmental conditions associated with impending rain, but is often triggered by either very high humidity, or in some cases low humidity and low temperatures.
The advertisement call is mostly made after dark and consists of a long series of repeated, very rapid “kek-kek-kek-kek-kek-kek” notes that can continue largely uninterrupted for several to many minutes. The sequence typically starts out relatively slowly then rapidly accelerates before stabilizing with very rapidly repeated keks. A momentary pause may ensue, with the slow to fast sequence repeated over between each pause.
Technical Reference:
Dodd (2013)
Online Photos:
Google
iNaturalist
Observation Methods:
The adults are best found by seeking calling males at the breeding sites or by riding roads on rainy night.
Download Video:
"MP4"
Download Video:
"MP4"
Download Video:
"MP4"
AmphibiaWeb Account
Distribution in North Carolina
Distribution Comments:
Hyla femoralis
is primarily found in the southeastern Coastal Plain from southeastern Virginia southward to southern Florida, then westward through southern Alabama and southern Mississippi to southeastern Louisiana. The range extends into the eastern Piedmont in a few areas, and a northern disjunct occurs in west-central Alabama and extreme eastern Mississippi. In North Carolina this species ranges throughout most of the Coastal Plain and extends into the extreme eastern Piedmont in a few areas.
Distribution Reference:
Beane et al. (2010), Dodd (2013)
County Map:
Clicking on a county returns the records for the species in that county.
GBIF
Global Distribution
Key Habitat Requirements
Habitat:
the Pine Woods Treefrog is strongly associated with southern pine forests of the Coastal Plain, but also can be found in mixed pine-hardwood associations. Although this species typically uses pine forests such as pine flatwoods and Longleaf Pine-wiregrass communities, it occasionally can be found in other habitats, including mesic and xeric hammocks, bottomland and slope forests, creek swamps, dome swamps, cypress bays, and sphagnum bogs (Dodd 2013). All of these communities have pine trees as significant components of the community.
The adults strongly prefer seasonal or semipermanent fish-free wetlands for breeding (e.g., Holbrook and Dorn 2015). These are typically found embedded within the forests or in close proximity to forest edges. Babbitt et al. (2006) found that occupancy rates were highest in breeding habitats in southern Florida that were within 50 m of forest edges, that lacked fish, and that were not strongly acidic. Examples of breeding sites listed by Dodd (2013) and Wright (1932) include temporary ponds, seasonal grassy depressions, woodland pools, cypress ponds, rain pools in fields, branch and creek swamps, Carolina Bays, depression marshes, cypress savannas, and cypress-gum ponds. Artificial habitats such as gravel pits and flooded ditches are also used.
In North Carolina this species is a denizen of the pine forests and woodlands of the Coastal Plain. It is common in naturally occurring Longleaf Pine savannas and flatwoods, and in Pond Pine Woodlands. It also occurs in silvicultural stands of Slash and Loblolly Pine, including pine plantations growing at the eastern edge of the Piedmont. Breeding sites that are used in North Carolina include depressional wetlands and other isolated or temporarily flooded pools, Carolina Bays, pools in swamp complexes, borrow pits, and roadside ditches.
Environmental and Physiological Tolerances:
Warner et al. (1991) grew tadpoles at three experimental acidities in outdoor artificial communities and found that survivorship and size at metamorphosis was lower at a pH of 4.3 relative to those grown in less acidic environments.
Adaptations to Natural Disturbances:
Natural populations of this species are associated with fire-maintained communities such as pine savannas, flatwoods, and oak scrub communities. Adults probably escape the once-typical ground fires of this region by climbing up into the canopy. Robertson et al. (2018) found no evidence of reduced genetic variability in local breeding populations at a fire-maintained ecosystem in Florida, and Gunzburger et al. (2010) found that populations recovered quickly after certain coastal areas had overwash that temporarily increased the salinity of breeding sites.
Biotic Relationships:
The adults usually select seasonal or semipermanent ponds for breeding. As with many species that use seasonal ponds, the larvae are palatable to fishes and other aquatic predators. Baber and Babbitt (2003) found that both native fishes in southern Florida and the introduced Walking Catfish (
Clarias batrachus
) will readily consume the tadpoles. These and other fishes sometimes colonize seasonal ponds in the region and severely impact tadpole populations. The seasonal drying of ponds eliminates fishes and other predators, and helps to return these sites to being productive habitats for anurans (Semlitsch et al. 1996).
The adults can potentially avoid fish-filled habitats that are death-traps to their future offspring by selectingly avoiding these. Rieger et al. (2004) examined the response of ovipositing females to the presence of varying numbers of the Eastern Mudminnow (
Umbra pygmaea
) in experimental pools and found strong avoidance behavior. Even a single small fish in a pool was sufficient to trigger strong avoidance behavior based on the number of eggs laid. Females appear to detect the presence of fish using chemical cues. In a companion study where 1-6 mudminnows were added per pool, larvae of
H. femoralis
suffered complete mortality when densities exceeded more than one fish per pool.
Odonates and other aquatic insects also feed on the larvae. Much like
H. chrysoscelis
, this species has evolved inducible defenses that reduce attack success. LaFiandra and Babbitt (2003) found that the presence of a large predatory dragonfly (
Anax junius
) triggered young tadpoles to development bodies and tails that were both deeper and shorter, along with tail fins with prominent orange coloration and bold black markings on the edges. The orange tail coloration and markings entice predators to strike the tail region rather than the more vulnerable head and body (McCollum and Van Buskirk 1996), and the broad tails improve maneuverability and likely increase the chance of escaping an attack. LaFiandra and Babbitt (2003) found that the induced responses were triggered both by alarm chemicals released from an injured tadpole and metabolites of digestion from the dragonfly larvae. Using a giant water bug (
Lethocerus americanus
) as a predator, McCoy (2007) found that tadpoles show graded responses and develop stronger defenses when the likelihood of being killed by a predator is higher.
The juveniles and adults appear to mostly rely on crypsis to avoid visually oriented predators. The brownish, bark-like dorsal pattern and tucked-in resting position provide camouflage when resting on pine limbs and bark (Dodd 2013). Individuals can also change color from light brown to blackish brown or green to better match their substrates. A variety of snakes eat the adults, including the North American Racer (
Coluber constrictor
), the Eastern Ratsnake (
Pantherophis alleghaniensis
), the Eastern Ribbonsnake
Thamnophis sauritus
, and the Common Gartersnake (
T. sirtalis
; Dodd 2013, Wright 1932).
See also Habitat Account for
General Coastal Plain Pine Forests and Woodlands
Life History and Autecology
Breeding and Courtship:
The adults are often active year-round except during cold bouts, but breeding tends to be concentrated in early summer. Thunderstorms and heavy rains can trigger the adults to leave their arboreal retreats and move towards the breeding sites (Duellman and Schwartz 1958). The males typically call from April to August or September (e.g., Dundee and Rossman 1989, Harper 1932, Mount 1975) but sometimes as early as February in north-central Florida and as late as October in southern Florida (Dodd 2013). Males in Georgia call from late-March to September (Jensen et al. 2008) and from March through October in the Carolinas and Virginia (Beane et al. 2010).
Breeding tends to be concentrated in late-spring and early summer, with occasional breeding bouts later in the year. Jensen et al. (2008) noted that most breeding occurs from April-June in Georgia, while Florida biologists have found tadpoles in ponds from mid-March through mid-December (Krysko et al. 2019). Egg laying in Virginia occurs from late May to early August (Mitchell 1986) and from May–July in North Carolina (AmphibiaWeb 2021). In the Sandhills of North Carolina, Wilbur (1997) only found tadpoles from mid- to late summer in water-filled tanks that were colonized by adults. Travis (1980) noted that breeding begins in May in this region, while Brandt (1936) found a mating pair on 1 May along the coast.
Males that arrive at the breeding sites mostly call at night, although a few may occasionally vocalize during the day. The calling sites are typically within 0-2 meters of the ground (Wright 1932). Common calling sites include saw palmetto leaves, shrubs and trees in or near the water's edge, bare ground, moss mats or tufts of vegetation at the pond margin, and downed logs, floating debris, and emergent vegetation in the pond proper. There can often be 50 or more males calling on a given night and the noise can be deafening (Dodd 2013). Pairing involves axillary amplexus, and egg laying and fertilization typically occurs within a few hours after amplexus (Wright 1932).
Merricks (2014) analyzed the advertisement calls of males and found that variation in the pulse rate is extraordinarily high relative to other hylid frogs. Moreover, the within-male pulse rate has a bimodal distribution within a single calling bout. Variation in the pulse rate becomes more pronounced as the chorus density increases. Other experiments that were conducted indicate that female
H. femoralis
show no selectivity based on the degree of variation in pulse rates, at least when they fall within the typical range of variability for this species.
Gerhardt (1974a) found that female
H. femoralis
can discriminate among potential mates based on mating call characteristics, and signal timing may play a strong role in female mate choice. Females show strong preferences for males that call immediately after a leading male stops calling, while those that strongly overlapped a calling male are less preferred (Merricks 2014).
Reproductive Mode:
Females deposit their eggs in monolayers to form rafts of floating eggs. The rafts tend to be roughly circular to elliptical in shape and can vary from 1.5–10 cm × 10–18 cm wide (Dodd 2013, Wright 1932). A female will normally deposit several to many rafts in a given night. Rieger et al. (2004) reported that individuals in Virginia typically deposit an average of around 700 eggs per breeding event, with the rafts containing around 50-200 eggs each. Wright (1932) found rafts with up to 150-200 eggs in Georgia, but commonly with many fewer. The freshly laid eggs are brown above and yellow below and are surrounded by two envelopes. The ova vary from 0.8–1.2 mm in diameter. The inner envelope varies from 1.4–2 mm in diameter, and the outer 4–8 mm in diameter (Dodd 2013). Wright (1932) observed several instances of complete reproductive failure in Georgia when shallow breeding sites dried shortly after the eggs were laid.
Clutch sizes are rather poorly documented. Delis (2001; cited in Dodd 2013) reported clutches of from 205–1,948 ova (mean = 924) in Florida specimens and clutch size was not correlated with the female's SUL. Sixty-five females from Virginia had from 701-2,086 ova (mean = 1,290; Mitchell and Pague 2014). Rieger et al. (2004) found that females lay around 700 eggs during a single breeding bout, but it is uncertain if they return to breed again later in the season. The embryos develop very quickly and can hatch within 1-3 days (Rieger et al. 2004, Wright 1932).
Aquatic Life History:
The hatchlings grow rapidly in the warm, shallow, seasonal ponds that are frequently used as breeding sites. Rieger et al. (2004) noted that the larval period lasts only about a month at their study site in Virginia, while Wright (1932) estimated that 40-70 days is most likely for populations in southeastern Georgia. Tadpoles that were grown in experimental ponds by Warner et al. (1991) under different combinations of pH and larval densities required an average of 36-51 days to metamorphose.
Very little reliable data are available on growth rates, survival, and length of the larval period in natural ponds, primarily because of staggered breeding and the difficulty of recognizing different age cohorts. These undoubtedly vary between ponds and years depending on ambient conditions such as the seasonal time of breeding, water temperatures, the quality and quantity of food, pond hydroperiods, the extent of both intra- and interspecific competition, and the types and densities of predators in the ponds. Crowding can result in slower growth rates, longer larval periods, lower survival, and smaller sizes at metamorphosis and can potentially act as a density-dependent mechanism to regulate local population sizes (Warner et al. 1991). The fully grown tadpoles can reach 33-36 mm TL, and very limited data suggest that the metamorphs are around 11-14 mm SUL (Dodd 2013, Wright 1932).
Breeding sites normally have an array of aquatic predators that feed on the larvae. Like several other hylids, the larvae have inducible defenses that reduce predation risk (see biotic interactions above). These tend to be costly to develop, and only do so when predators such as dragonfly larvae are present in the ponds.
Terrestrial Life History:
We have much to learn about the terrestrial ecology of this species. In southeastern Georgia, the young metamorphs from the primary breeding season in early to mid-summer grow rapidly and most appear to reach sexual maturity by the following spring based on reports of SUL (Wright 1932). Most males likely breed during their first year of life, with perhaps some females taking an additional year before laying their first clutch of eggs. Individuals from very late-breeding cohorts may not be able to breed during their first year.
The young metamorphs are often found close to wetlands on low-lying vegetation and it is uncertain to what extent they are arboreal. They appear to ultimately move substantial distances from the breeding sites. Dodd (1996) collected individuals in Florida that were an average of 317 m (range = 42–815 m) from the nearest breeding sites. The adults are strongly arboreal except during their migrations to and from the breeding sites, and many aspects of their arboreal life history such as diet, home range, and territoriality are largely unknown. They can tolerate the hot and dry conditions associated with living in trees by coating their skin with a waterproofing layer of mucous and lipids that retard water loss (Barbeau and Lillywhite 2005, Dodd 2013, Wygoda 1984). They also take cover in microhabitats such as loose bark, pine stumps and tree holes. Individuals in northern populations probably overwinter on the ground like
H. chrysoscelis
, but this has not been documented.
The diet is poorly documented, but the limited information that is available suggests that individuals are gape-limited generalists. Seventeen specimens from southern Florida that were examined by Duellman and Schwartz (1958) had eaten grasshoppers, a cricket, beetles, a caddisfly, ants (Formicidae), wasps (Vespidae), and a jumping spider. Three additional specimens had empty guts. These were presumably adults that were at or near the breeding ponds.
General Ecology
Population Ecology:
Local populations are centered around the breeding sites. Estimates of local population sizes are largely lacking because the adults easily trespass across drift fences that are used to capture many amphibians. Greenberg and Tanner (2005) collected 488 adults at a cluster of 8 breeding ponds in Florida during a multiyear survey, but this undoubtedly is a gross underestimate of the actual numbers of breeding adults.
Robertson et al. (2018) analyzed genetic variation in 17 breeding aggregations within a 2,100-ha study area in Florida that had been exposed to different fire regimes. They found that local breeding aggregates had high levels of genetic connectivity throughout the study area and behaved as a single panmictic population. Genetic variation was high in local populations and there was little evidence that either recent or frequent fires reduce genetic diversity or restrict connectivity among pond populations.
Community Ecology:
The Pine Woods Treefrog often shares breeding ponds with other anuran larvae, but ecological interactions between these species have largely been confined to studies in laboratory aquaria or artificial ponds. Wilbur (1980) grew hatchlings of
H. femoralis
and
H. gratiosa
at different densities in laboratory aquaria and found that
H. gratiosa
appeared to be a superior competitor. Warner et al. (1993) examined the interactions of pH and tadpole density on tadpoles of
H. femoralis
and
H. gratiosa
in outdoor tanks. Crowding of
H. femoralis
resulted in lower growth rates, higher mortality, and a smaller size at metamorphosis. Larvae also performed poorly at the lowest pH (4.3). The authors found that prior use of the tanks by
H. gratiosa
earlier in the year actually enhanced the performance of
H. femoralis
.
Adverse Environmental Impacts
Habitat Loss:
This species was a characteristic faunal element of the extensive Longleaf Pine ecosystems that blanketed the southeastern Coastal Plain in colonial times. It appears to be well adapted to tolerating the periodic surface fires that maintained these forests (Langford et al. 2007). Most of the Longleaf Pine forests were subsequently destroyed (Outcalt and Sheffield 1996), and it is uncertain to what extent populations of
H. femoralis
were adversely affected. The widespread loss of seasonal ponds in the Coastal Plain has undoubtedly eliminated many populations. In addition, the loss or degradation of forested habitats has been widespread due to urbanization, clearcutting, and the conversion of forested land to pastures, agricultural fields, and pine plantations (Dodd 2013, Haggerty et al. 2019, O’Neill 1995, Pieterson et al. 2006). Despite the ever-increasing impact of humans on natural systems, the Pine Woods Treefrog is still common in many areas -- at least by modern standards (Dodd 2013, Krysko et al. 2019).
Habitat Fragmentation:
Populations appear to tolerate habitat fragmentation to some extent so long as extensive tracts of pine forests remain within close distances to the breeding sites. This species uses roadside ditches and flooded fields in many areas, which has helped to ameliorate the loss of natural wetlands.
Status in North Carolina
NHP State Rank:
S5
Global Rank:
G5
Status Comments:
Despite widespread historical losses due to the loss or degradation of wetlands and the loss of previously forested areas in the Coastal Plain,
H. femoralis
show no evidence of marked ongoing declines within the state.
Stewardship:
This species does best in Longleaf Pine communities. Fire suppression that allows pines to be replaced by hardwoods can result in the loss or decline of populations. Seasonal, fish-free breeding sites provide optimal habitats for the tadpoles and are an essential feature for maintaining healthy local populations. Prescribed burns are critical in many areas for sustaining healthy local populations.
Recording Gallery for
Hyla femoralis
- Pine Woods Treefrog
Your browser does not support the audio element.
2022-05-24. Beaufort Co. Jim Petranka and Becky Elkin - Tree calls issued from tops of Loblolly Pines.
Your browser does not support the audio element.
2022-05-25. Beaufort Co. Jim Petranka and Becky Elkin - advertisement calls from a vegetated ditch with H. chrysoscelis.
Your browser does not support the audio element.
2024-05-27. Harnett Co. Paul Hart -
Photo Gallery for
Hyla femoralis
- Pine Woods Treefrog
12 photos are shown.
Recorded by: Mark Shields
Onslow Co.
Recorded by: Jim Petranka and Becky Elkin
Brunswick Co.
Recorded by: Jim Petranka and Becky Elkin
Brunswick Co.
Comment: A view of the rear thighs with bright yellow-orange spotting.
Recorded by: Steve Hall, Jim Petranka, and Bo Sullivan
Richmond Co.
Comment: test
Recorded by: Steve Hall
Columbus Co.
Recorded by: Steve Hall
Columbus Co.
Recorded by: Steve Hall
Columbus Co.
Recorded by: Mark Shields
Onslow Co.
Recorded by: Stephanie Chavez
New Hanover Co.
Recorded by: Steve Hall
Brunswick Co.
Recorded by: Steve Hall
Brunswick Co.
Recorded by: Steve Hall
Hoke Co.

 »
»