Taxonomy
Class: AmphibiaOrder: AnuraFamily: HylidaeSubfamily: Hylinae
Taxonomic Comments: Pseudacris ferianum is a member of the trilling chorus frog group, which is a complex of species that have had a confusing taxonomic history. Lemmon et al. (2007) conducted a comprehensive molecular analysis and made major revisions in historically recognized species. They also documented two major clades within P. ferianum , with North Carolina populations belonging to the "coastal" clade.Species Comments: The genus Pseudacris has proved to be a taxonomically challenging group with numerous names of species and subspecies applied to forms in the eastern US that no longer fit the current taxonomy that is based on molecular analyses. Dodd (2013) noted that there is considerable taxonomic confusion in the published literature, so one has be careful when reading older papers that seem to apply to a species such as P. triseriata , but actually do not based on known ranges of the currently recognized species.
Identification
Description: The adults are small frogs that have slightly enlarged toe pads and granular skin. The dorsal ground color is commonly tan or light to medium brown, but can be other shades such as gray, dark brown, or greenish brown. Many specimens have three brownish longitudinal stripes down the back that are darker than the prevailing ground color. In some specimens these are broken into a series of linear spots or blotches, while in others they may be missing or nearly so, particularly in Coastal Plain populations from South Carolina (Dodd 2013). Many specimens also have a dark triangular mark between the eyes, but it is commonly missing on weakly marked specimens. A dark band is present on each side of the head that begins near the nostril and widens as it passes through the eyes to the shoulder region. In some specimens it may continue beyond the shoulder region where it tends to break up before reaching the groin area. The upper lip has a white stripe that contrast with the darker band above. The belly and chest are white to cream-colored and some specimens may have darker flecks or spotting that are most prominent on the chest. Vocalizations: The advertisement calls consist of slow trills, sounding like a "crreeek" that has been likened to running one's finger along the tines of a pocket comb. Since each such call is frequency modulated, increasing in pitch from start to finish, the comb analogy works if direction of the finger's movements are from thick tines at the beginning to thinner tines towards the end. At 60 F, the dominant pitch rises from 2 kHz at the start to 3.5 at the end. Some modulation has also been noted in the pulse rate within a call: the notes tend to be closer together at the beginning of the call compared with the end of the call (Dodd 2013, Thompson and Martof 1957). We have noticed that pulses may be doubled at the beginning but single towards the end, but with the pulse rate otherwise fairly uniform. The call rates are temperature dependent, but typically range from 0.5-1.25 per second. They are repeated about every second or so. In addition to the advertisement calls, males produce a harsh release call that is issued if one male amplexes a second (Martof and Thompson 1958).P. nigrita ) in a contact zone at the junction of the Piedmont and Coastal Plain, and with Brimley’s Chorus Frog (P. brimleyi ) in South Carolina north into Virginia. In areas of overlap with (P. nigrita ), the vocalizations of P. feriarum have a faster pulse rate relative to populations outside of the zone of overlap. This appears to reflect reproductive character displacement that reduces breeding mistakes with congenerics (e.g., Lemmon 2009, Malone et al. 2014). Hybrids from experimental crosses of the two species have much lower fitness relative to the offspring from homospecific crosses (Lemmon and Lemmon 2010).Technical Reference: Dodd (2013)Online Photos: Google iNaturalist Observation Methods: The adults are most easily observed as they call during the day and night from seasonal wetlands. They are also commonly encountered on rainy nights during the breeding season.
Download Video:
"MP4"
AmphibiaWeb Account
Distribution in North Carolina
Distribution Comments: The Upland Chorus Frog is found in the eastern US from central Pennsylvania southward in a sweeping arc through Virginia and the Carolinas into Georgia, Alabama, the Florida Panhandle, and northeastern Mississippi. From there the range extends northward through most of Tennessee, extreme northeastern Arkansas, and southeastern Missouri before terminating in southern Illinois and extreme southwestern and south-central Kentucky (Cambridge 2018). This species is generally uncommon or absent from most of the southern Appalachians. A disjunct population occurs near the coast in South Carolina in Berkeley, Dorchester, and Charleston counties (Schwartz 1957, Lemmon et al. 2007).P. feriarum is primarily found in the Piedmont and Upper Coastal Plain, although there are a few records from farther east along the major river drainages. Populations are common and widespread in the eastern and central Piedmont, and become progressively scattered farther west towards the Blue Ridge. This species is absent or very rare in the Blue Ridge, although fringing populations occur in Tennessee very close to the state line. A recent record was obtained from Madison County.Distribution Reference: Beane et al. (2010), Dodd (2013), Lemmon et al. (2007)County Map: Clicking on a county returns the records for the species in that county.
GBIF Global Distribution
Key Habitat Requirements
Habitat: The adults are strongly affiliated with moist deciduous forests. Bottomland forests, swamp forests, or mesic hardwoods are used by the adults outside of the breeding season. Breeding typically occurs in fish-free seasonal wetlands that are either embedded in forests or are in open habitats nearby. Sites that are in open areas are often shallow and vegetated with rushes, sedges, grasses or other aquatic vegetation. Some of the commonly used habitats include woodland vernal pools, marshes, and seasonal pools in cypress/gum swamps. This species commonly breeds in disturbed habitats and will use small pools and puddles in roadways, shallow ditches, as well as temporary pools in pastures or croplands. Biotic Relationships: The adults appear to almost always breed in fish-free habitats and the larvae are palatable to native fishes, amphibian larvae, and aquatic invertebrates. Stanback (2010) showed that the small larvae were quickly eliminated from experimental pools with the Eastern Mosquitofish (Gambusia holbrooki ). The larvae are also readily eaten by odonates and giant water bugs, and will reduce movements when exposed to chemical cues from predators that were fed tadpoles (Haislip et al. 2012). Wood Frog tadpoles can be important predators on the eggs -- and to a lesser extent -- the hatchlings (Sours and Petranka 2007). Other known predators include Ambystoma larvae (Stemp et al. 2021), the Eastern Newt, water snakes (Nerodia ), and garter snakes (Thamnophis ; Dodd 2013). Dodd and Cupp (1978) reported that Upland Chorus Frog larvae that are approaching metamorphosis have an immobility response where they draw their limbs into the body and refrain from any movement if approached.
Life History and Autecology
Breeding and Courtship: The Upland Chorus Frog is an early seasonal breeder, with calling and breeding occurring from winter through early spring. Dodd (2013) noted that the adults begin to congregate near the breeding sites in the fall and remain there throughout the winter. As such, there are no strong breeding migrations to the ponds shortly before chorusing begins as seen in many seasonal pond breeders. The males may occasionally call in November, but calling in earnest generally does not begin until December or January in the southernmost portions of the range, and not until February or later in the northern portions of the range. Dodd (2013) summarized the reported breeding dates for several sites throughout the range. Examples include December to March in Alabama, January to March in Mississippi, January to April in Tennessee, and March to April in West Virginia. Breeding in North Carolina on rare occasions can occur as early as December, but more commonly occurs from January to April depending on the location (Alexander 1966, Murphy 1963, Steelman and Dorcas 2010). February and March appear to be the peak months at most sites. Reproductive Mode: Each female lays a series of small egg masses. These typically are cylindrically shaped, about 2-2.5 cm wide and have around 10-100 eggs per mass. The jelly has a loose consistency, and the masses are attached to submerged support structures such as twigs, branches, and the stems and leaves of sedges and grasses. Estimates for the number of eggs per mass include 10-40 from Alabama (Brown 1956), 8-35 from Georgia (Martof and Thompson 1958), and 30-50 eggs from Tennessee (Britson and Kissell 1996). Alexander (1966) reported extremes of 4-159 eggs for North Carolina masses. Four masses that were observed by Jim Petranka in Madison Co. contained from 62-101 developing embryos (mean = 78). When depositing a mass, the female straddles and clasps a stem or other support structure. She then arches her back to bring her cloaca nearly into contact with the supporting structure and the vent of the male. As the eggs are extruded, the male fertilizes them (Martof and Thompson 1958). Aquatic Life History: The larvae are detritus feeders that feed on bottom muck and decaying vegetation. They use the jaws and teeth to rasp detritus and other potential food items from the substrate, which is then picked up by a stream of mucous and carried into the esophagus (Dodd 2013). They will also feed on dead animal remains. The larvae are solitary feeders that tend to be uniformly distributed throughout a pool. The quality of food resources can affect larval growth and development. Britson and Kissell (1996) fed larvae different food types and found that larvae that were fed pollen from pine and oak trees performed poorly relative to those fed algae or commercial animal food. Animals that fed on pollen grew slower and were less likely to initiate metamorphosis, perhaps due to a lack of iodine in oak pollen. P. feriarum . This trade-off may explain why the breeding season is relatively long for this species, since early breeders may be favored in wet years, but not in years with premature pond drying. Terrestrial Life History: Studies by Alexander (1966) and Murphy (1963) in North Carolina indicate that the young metamorphs remain in the immediate vicinity of ponds for several weeks before moving to nearby forests (Dodd 2013). Alexander (1966) found that the young juveniles began dispersing away from the ponds after reaching about 20 mm SUL. Dispersal required several weeks to complete and most individuals moved to a nearby swamp forest where they tended to hide in very thick vegetation. Murphy (1963) found that most movements occur during or immediately following rainfall, and Jensen et al. (2008) reported that individuals move as far as 200 m from the breeding sites.
General Ecology
Population Ecology: The Upland Chorus Frog is organized as small local populations that are centered around local breeding sites. We have little data on population organization in this species. The juveniles appear capable of dispersing several hundred meters from the breeding sites, but we need more information on breeding site fidelity and the extent to which populations have metapopulation structure. Community Ecology: Pseudacris feriarum often shares breeding ponds with several other early breeding amphibians, but we have little information on competitive interactions with other species. Wood Frog tadpoles can adversely affect P. feriarum by consuming their eggs and competing with the tadpoles (Sours and Petranka 2007). Predators undoubtedly consume large numbers of tadpoles. Chemical cues from predators can trigger the tadpoles to reduce their activity and feeding levels (Haislip et al. 2012), which in turn could increase their chances of becoming trapped in drying ponds. Jensen et al. (2008) reported that this species usually breeds in open canopy habitats but switches to forested gum or cypress swamp habitats when they are sympatric with P. nigrita . This suggests that P. feriarum might be competitively excluded from their preferred breeding sites.
Adverse Environmental Impacts
Habitat Loss: As with most amphibians, habitat loss is one of the greatest threats to species. The Upland Chorus Frog undoubtedly suffered significant losses of populations after European colonization as large areas of deciduous forests were cleared and seasonal wetlands were filled. Populations today have compensated for the loss of natural wetlands by shifting breeding to roadside ditches, flooded fields, and other artificial breeding sites. However, they have not been able to compensate for the widespread loss of bottomland forests from throughout the range of this species. Habitat Fragmentation: Populations appear to tolerate habitat fragmentation reasonably well so long as breeding sites are available within close proximity to wet or mesic deciduous forest. Additional information is need on the extent to which extant populations are interconnected via dispersal between local populations.
Status in North Carolina
NHP State Rank: S5Global Rank: G5Environmental Threats: Deforestation, urbanization, and the filling of seasonal wetlands are the greatest threats to this species. Populations have adapted to fragmented landscapes by shifting to using artificial habitats such as roadside ditches and flooded agricultural fields that are in close proximity to floodplain or mesic forests. This has compensated for the loss of natural seasonal wetlands and has allowed populations to stay relatively stable despite significant impacts associated with population growth in North Carolina. Road kill is a significant cause of mortality in some areas. Status Comments: Populations in North Carolina do not show any evidence of recent widespread declines and appear to be stable. Stewardship: Local populations are best maintained by have clusters of seasonal ponds in close proximity to wet or mesic deciduous forests. This species does not use permanent ponds with fishes, and does best in habitats with sunny or partially shaded breeding sites.
Recording Gallery for Pseudacris feriarum - Upland Chorus Frog
Your browser does not support the audio element.
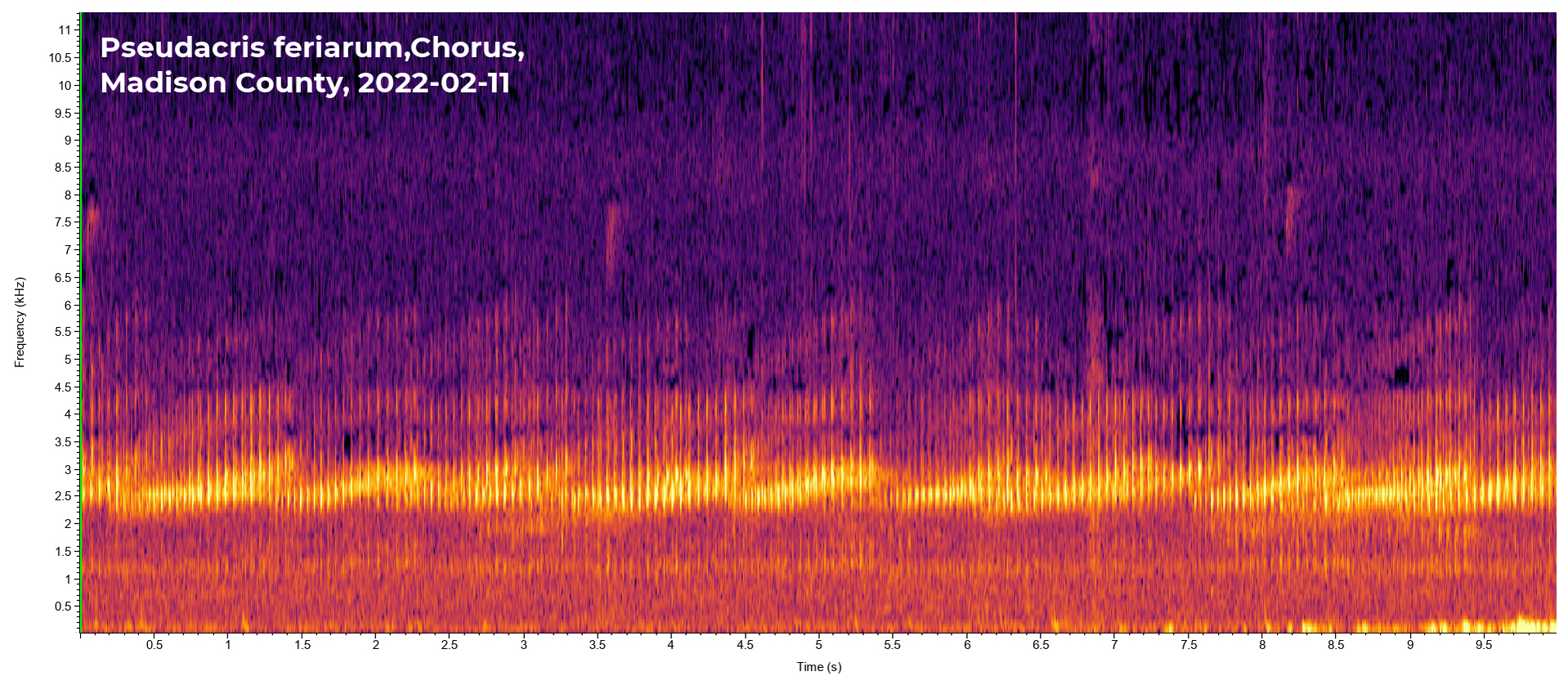
2022-02-11. Madison Co. Jim Petranka and Becky Elkin - Males were calling from a ditch along RR track; 1,301'
Your browser does not support the audio element.
2022-02-11. Orange Co. Steve Hall and Savannah Hall -
Your browser does not support the audio element.
2022-03-02. Madison Co. Jim Petranka and Becky Elkin - Around 10 adults were calling around noon from a shallow ditch adjoining an agricultural field. Eggs of Pseudacris, A. maculatum, and L. sylvaticus were in the ditch. Air temp. = 70F.
Your browser does not support the audio element.
2022-03-02. Madison Co. Jim Petranka and Becky Elkin - Two adults calling from a woodland pond; air temp. = 70F
Your browser does not support the audio element.
2022-02-18. Orange Co. John Petranka -
Your browser does not support the audio element.
2024-02-13. Alexander Co. Guy McGrane -
Photo Gallery for Pseudacris feriarum - Upland Chorus Frog 13 photos are shown. Recorded by: David George Recorded by: Steve Hall and Savannah Hall Recorded by: Rob Van Epps Recorded by: Jim Petranka and Becky Elkin Recorded by: Jim Petranka and Becky Elkin Recorded by: Steve Hall Recorded by: Travis McLain Recorded by: Owen McConnell Recorded by: Steve Hall and Harry LeGrand Recorded by: Steve Hall and Harry LeGrand Recorded by: Steve Hall Recorded by: Steve Hall Recorded by: Steve Hall

 »
»