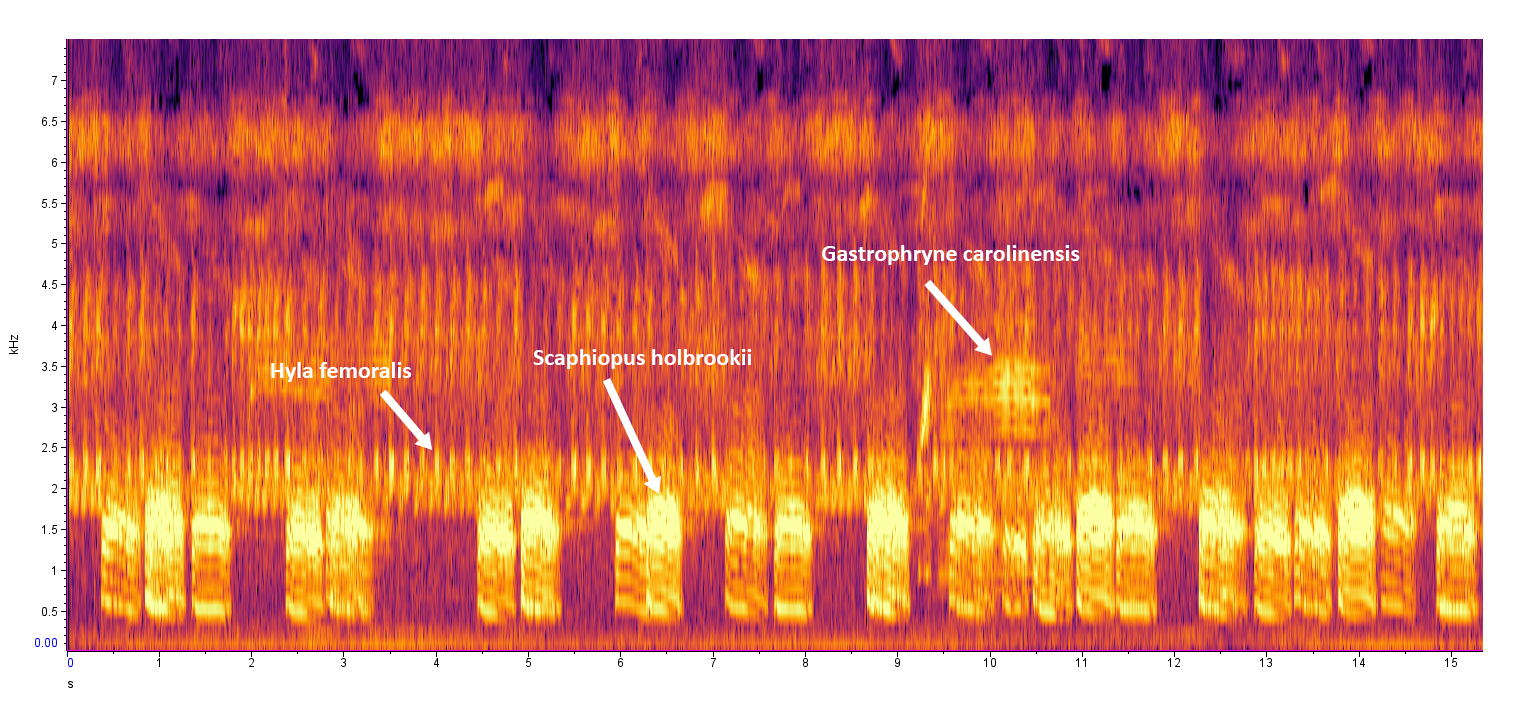
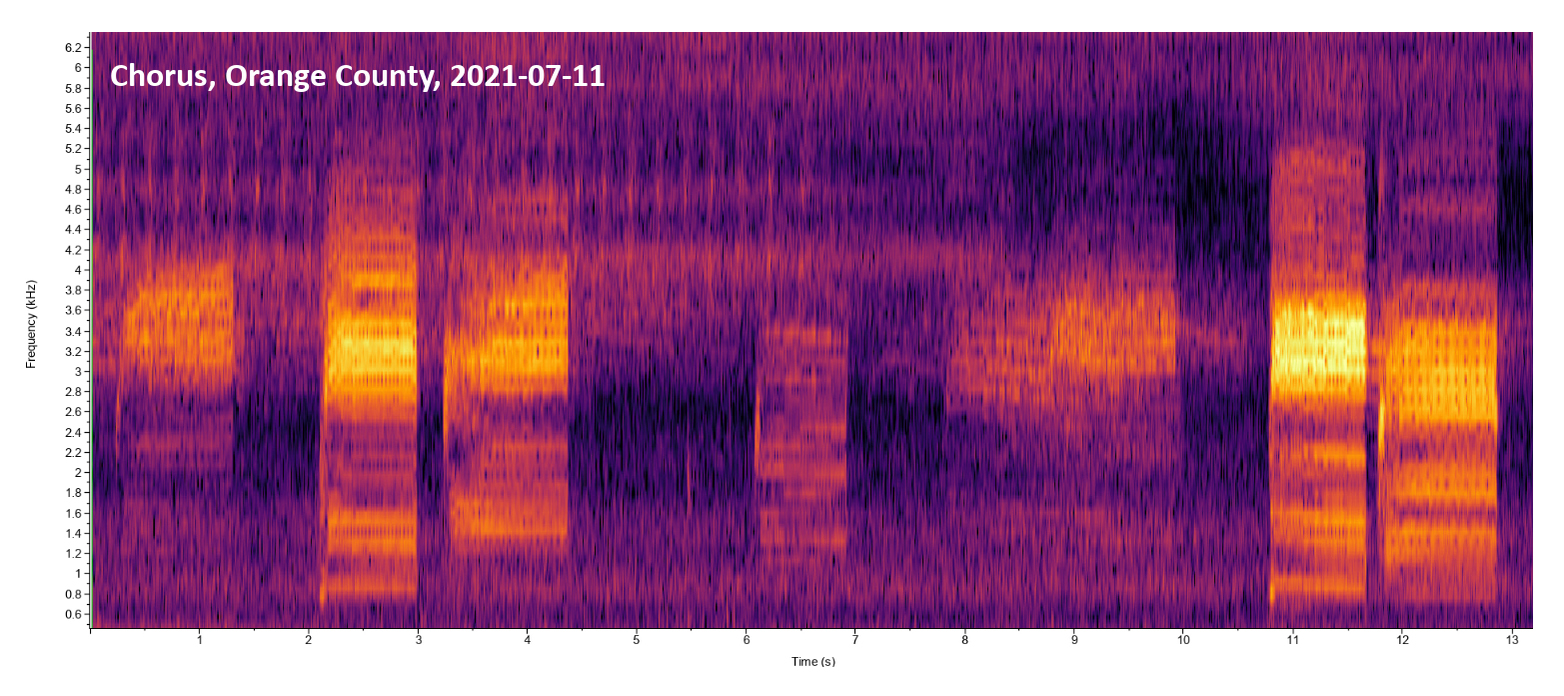
|
|
|
|
|
|
|
|
Photo Gallery for Gastrophryne carolinensis - Eastern Narrow-mouthed Toad
| 37 photos are available. Only the most recent 30 are shown.
|
 | Recorded by: Mark Shields
Onslow Co. |  | Recorded by: Mark Shields
Onslow Co. |
 | Recorded by: Mark Shields
Onslow Co. |  | Recorded by: Andrew W. Jones, Atley Elliott
Polk Co. |
 | Recorded by: Andrew W. Jones, Atley Elliott
Polk Co. |  | Recorded by: Andrew W. Jones
Polk Co. |
 | Recorded by: Andrew W. Jones
Polk Co.
Comment: Eggs that were deposited in shallow water. |  | Recorded by: R. Newman
Carteret Co. |
 | Recorded by: A. Ledford
Wake Co. |  | Recorded by: Steve Hall and Savannah Hall
Orange Co. |
 | Recorded by: Morgan Freese
New Hanover Co. |  | Recorded by: Hunter Phillips
Onslow Co. |
 | Recorded by: Morgan Freese
Dare Co. |  | Recorded by: Morgan Freese
Dare Co. |
 | Recorded by: j.wyche
Gates Co. |  | Recorded by: j.wyche
Gates Co. |
 | Recorded by: j.wyche
Gates Co. |  | Recorded by: Morgan Freese
Durham Co. |
 | Recorded by: G. Schneider, J. Murvine
Stanly Co. |  | Recorded by: G. Schneider, J. Murvine
Stanly Co. |
 | Recorded by: F. Williams, S. Williams
Gates Co. |  | Recorded by: j.wyche
Gates Co. |
 | Recorded by: j.wyche
Gates Co. |  | Recorded by: J. Wyche5
Gates Co. |
 | Recorded by: J. Wyche5
Gates Co. |  | Recorded by: W. Stanley
Cumberland Co. |
 | Recorded by: W. Stanley
Cumberland Co. |  | Recorded by: J. Wyche
Gates Co. |
 | Recorded by: Jane Wyche
Gates Co. |  | Recorded by: Jane Wyche
Gates Co. |
|

 »
»