Amphibians of North Carolina
Home Page
Recent Entries
Recent Account Updates
General Search
Submit a Public Record
References
Maps
Checklist
NC Biodiversity Project
Comments
Scientific Name:
Common Name:
Family (Alpha):
AMBYSTOMATIDAE
AMPHIUMIDAE
BUFONIDAE
CRYPTOBRANCHIDAE
HYLIDAE
MICROHYLIDAE
PLETHODONTIDAE
PROTEIDAE
RANIDAE
SALAMANDRIDAE
SCAPHIOPODIDAE
SIRENIDAE
«
Home
»
Sole representative of Scaphiopodidae in NC
Scaphiopus holbrookii
- Eastern Spadefoot
Taxonomy
Class:
Amphibia
Order:
Anura
Family:
Scaphiopodidae
Taxonomic Comments:
Species Comments:
This species has a noxious, musty or peppery smell and produces skin secretions that can cause the mucous membranes to burn and the eyes to water.
Identification
Description:
The Eastern Spadefoot is a medium-sized frog with a blunt snout, vertical pupils, and a black spade-like projection on each hind foot. The dorsal ground color varies from blackish to grayish or olive yellow, and there are usually two light yellow lines that run down the back. In many specimens these form a lyre-like pattern where they begin near the back of the eyes, converge slightly towards mid-body, then bow outward towards the back legs before reuniting near the urostyle. However, the paired lines can be fragmented, greatly reduced in size, or even missing in some specimens. A second and often less conspicuous line is usually present along each side. The dorsum is slightly rugose and may have a few very small warts, while the venter is white to grayish white and unmarked.
This species lacks cranial crests but has rounded parotoid glands that are otherwise similar to those of our native toads. In addition to the parotoid glands, there are two distinct pectoral glands on the undersides just below the front limb insertions. These appear as raised bumps and are anatomically similar to the parotoid glands of toads (Jacob et al. 1985). The adults vary from 45–78 mm SUL. Examples of average lengths that have been reported for males and females include 59 mm and 61 mm in Connecticut, 58 mm and 53 mm in northern Florida, and 52 and 48 mm in Key West. Males show a weak tendency to be slightly larger than the females (Dodd 2013).
The tadpole is dark bronze to brownish above with small orange spots and a round body when viewed from above. The eyes are very narrowly set apart, and the head is wide relative to the body width. The tail is short and rounded, and has unmarked dorsal and ventral fins (Dodd 2013).
Vocalizations:
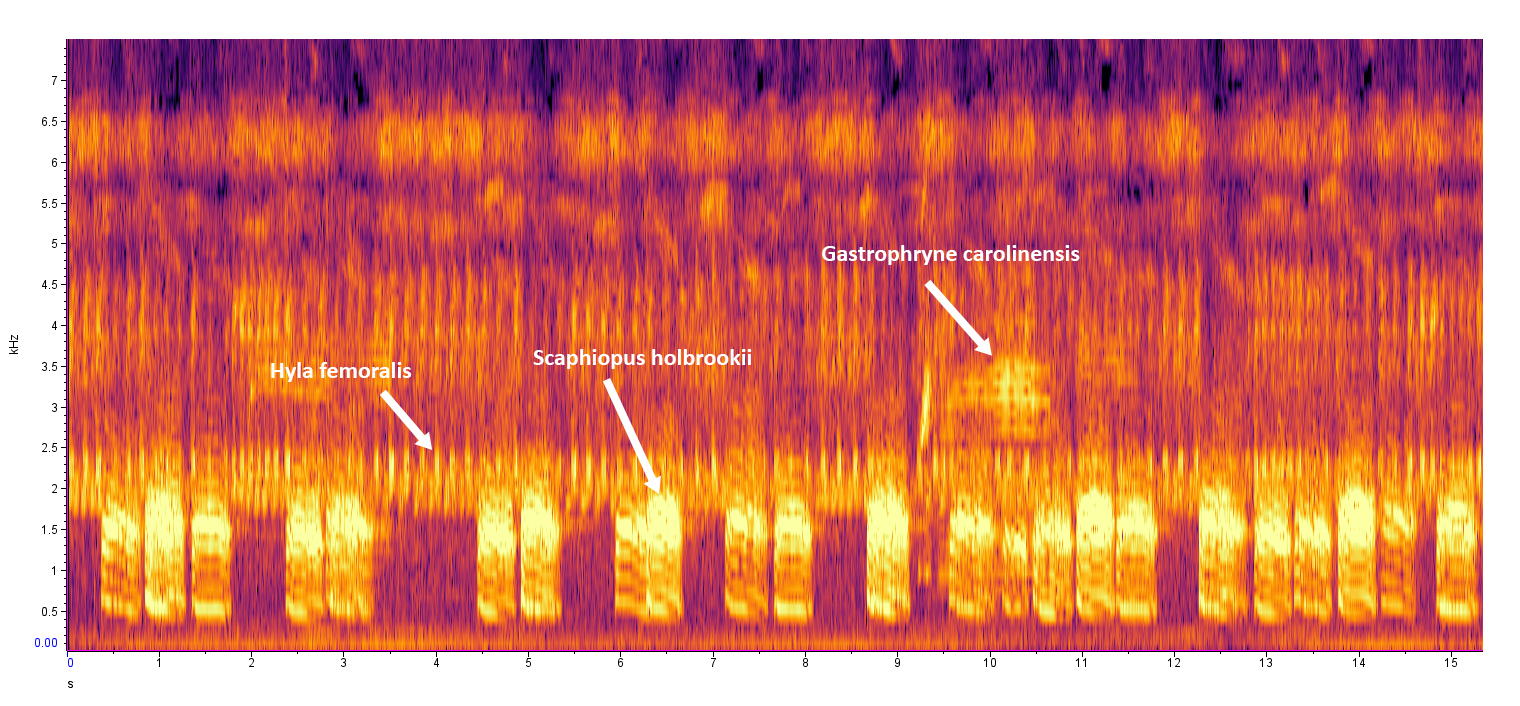
The males have a distinctive loud, harsh, nasally call that is difficult to describe. It has been likened to the second syllable in a long drawn-out meow of a Siamese cat (“owww, owww”) or the “waagh” or “waaank” call of a young crow (Dodd 2013). The call last for about one-half of a second and is repeated every 4–6 seconds.
Online Photos:
Google
iNaturalist
Observation Methods:
The adults spend most of their time in underground burrows and are best found by either driving roads on rainy nights or listening for calling males during bouts of heavy rainfall.
Download Video:
"MP4"
AmphibiaWeb Account
Distribution in North Carolina
Distribution Comments:
Scaphiopus holbrookii
is restricted to the eastern United States where it occurs from Massachusetts southward along the Eastern Seaboard to southern Florida and the Florida Keys. The range extends from Georgia and Florida westward across Alabama and Mississippi before ending in southeastern Louisiana. From there the range extends northward through eastern Arkansas, Tennessee and Kentucky to southeastern Missouri, southern Illinois, southern Indiana, southern Ohio, and western West Virginia. Populations are relatively common in the Coastal Plain, but are patchily distributed elsewhere where they tend to be commonly found in river valleys with sandy, alluvial soils. The species is generally absent from the Appalachian and Allegheny Mountains, but a few scattered populations are known from mountainous regions in the lower valley floodplains (Dodd 2013).
In North Carolina, populations are most common in the southern half of the Coastal Plain and eastern Piedmont, particularly in the Sandhills and coastal pinelands. However, numerous other populations have been documented in other areas of the Coastal Plain and on the Outer Banks. We also have a few widely scattered records from the western Piedmont and southwestern mountains, although several of these are historical.
Distribution Reference:
Beane et al. (2010); Dodd (2013)
County Map:
Clicking on a county returns the records for the species in that county.
GBIF
Global Distribution
Key Habitat Requirements
Habitat:
The juveniles and adults are found in a wide assortment of terrestrial habitats, the key being the presence of friable soils that allow individuals to dig with the hind legs and bury themselves. Sites with sandy soils or sandy loams are ideal. Most populations are found on the Coastal Plain at sites with well-drained soils and shallow, seasonal wetlands for breeding. They inhabit both mesic and dry sites, but are rarely found where the soil is permanently saturated. Populations that are found outside of the Coastal Plain commonly use floodplains and river terraces with friable alluvial deposits. Habitats that are used throughout the range include pine flatwoods, scrub and sandhill communities, xeric and mesophytic hammocks, upland hardwoods, and steephead ravines (Dodd 2013). Population are regularly found also in disturbed habitats such as suburban developments, agricultural fields, rural road corridors, pastures, and managed pine forests so long as the soils are friable enough to allow digging and burrowing.
The larval period is very brief and the adults typically select seasonal, fish-free ponds and flooded depressions for breeding. Breeding normally occurs during or immediately following heavy rain events that fill seasonal ponds or create shallow flooded depressions in both natural or disturbed settings. Common breeding sites include natural seasonal wetlands in either forested or non-forested habitats, floodplain depressions, Carolina bays, flooded agricultural fields, flooded pastures, ditches, borrow pits, and retention ponds. Larger and deeper sites are occasionally used, but the eggs are deposited in the shallowest section of these wetlands. In North Carolina, populations use most of the habitats listed above.
Adaptations to Natural Disturbances:
Populations appear to tolerate both periodic fires and disturbance from hurricanes and other major storms.
Biotic Relationships:
The adults normally breed in shallow ephemeral ponds that have relatively few aquatic predators, but sometime use less ephemeral sites with greater predator loads. The larvae appear to lack chemical defenses against fish (Kurzava and Morin 1998) and are almost always found in fish-free habitats. Dodd (2013) noted that the eggs are eaten by leeches, and the larvae by aquatic insects. Adults of the Eastern Newt are effective predators on the larvae in artificial communities, but it is uncertain how important this is in natural systems since newts generally prefer semipermanent or permanent habitats for breeding. The tadpoles use chemical cues to track predators and will rapidly flee from an area if an injured animal is placed among a group to simulate an attack by a predator (Richmond 1947).
The subadults and adults can reduce encounters with terrestrial predators by rapidly burrowing into the soil, This can take only a few seconds in sandy soils (Dodd 2013). If unable to dig, an individual will curl its body and tuck the chin downward, close its eyes, and draw the limbs and feet tightly inward and remain immobile (Dodd and Cupp 1978). This exposes the predator to noxious secretions on the dorsal surface of the body. Dodd (2013) summarized some of the known predators of the juveniles and adults. These include the Southern Toad (
Anaxyrus terrestris
), American Bullfrog (
Lithobates catesbeianus
), Southern Hognose Snake (
Heterodon simus
), Northern Water Snake (
Nerodia sipedon
), Banded Water Snake (
Nerodia fasciata
), North American Racer (
Coluber constrictor
), and Cottonmouth (
Agkistrodon piscivorus
). Bird predators include the Cattle Egret and European Starling as well as grackles, owls, and gulls, while mammalian predators include feral pigs, coyotes, raccoons and opossums. Beane et al. (2014) found that
S. holbrookii
is an important prey item of
H. simus
in North Carolina and surmised that the snakes may actively seek them out in their burrows.
See also Habitat Account for
General Mixed Habitats
Life History and Autecology
Breeding and Courtship:
The adults are explosive breeders and can breed at any time of the year in southern populations, and for most of the year in more northern populations except when in winter dormancy. Rather than having annual spermatogenic and oogenic cycles associated with seasonal breeding as seen in many temperate amphibians, the males retain viable sperm in their testes year-round and gravid females can be found throughout the year (Goldberg 2018, Hansen 1958). Breeding in North Carolina can occur during most months of the year. Records of calling or mating adults extend from February through October, while adults have been found moving on roads on rainy nights from late January through early December.
Breeding is triggered by heavy rains, and exceptionally heavy chorusing and breeding often occur is association with tropical storms and hurricanes. Males and females typically leave their burrows and arrive at the breeding sites after dark and within 6-18 hours after heavy, prolonged rainfall begins. The entire breeding period may last only a day or two, but sometimes for up to eight days depending on the duration of a rain event and weather conditions (Church et al. 2002, Dodd 2013, Pearson 1955). Breeding in Florida and other southern locales occurs year-round, and commonly during the winter and spring months when slow-moving cold fronts produce 3-4 inches of rain or more. Populations in the northern portion of the range are not active until the arrival of warmer spring weather and may not begin breeding until March or later (Goldberg 2018, Hansen 1958). More than one major breeding bout can occur in a local population during a single year. This likely reflects different cohorts of females breeding rather than individual females ovipositing multiple clutches (Ball 1936, Dodd 2013, Gosner and Black 1955). Adults may also skip breeding in certain years. Palis (2012), for example, tracked a sinkhole population in southern Illinois for 17 years and found that the adults engaged in 26 breeding bouts, with from 1–3 bouts per year. They did not call or mate during 5 of the 17 years.
The males call both during the day and night (Neill 1957) and females arrive shortly after the males begin calling. Driver (1936) even observed several excited males in Massachusetts calling from their underground burrows before digging out and moving to a nearby breeding site. Individuals call while floating in the water and arch their heads and bodies backwards when inflating the single vocal pouch. This allows them to the expand the vocal pouch above the water line and transmit more effectively through the air. Dodd (2013) noted that females are often amplexed on land as they make their way toward the breeding site. In addition, many males at a breeding site remain silent and simply intercept and clasp females that are swimming in the ponds. Amplexus is inguinal, with the male grasping the female immediately in front of the insertion of the hind legs. Once amplexed, the female selects an oviposition site in shallow water.
Reproductive Mode:
Each female attaches several groups of loosely clumped eggs to leaves, stems, submerged vegetation, or other support structures. The eggs are deposited in cylinders or bands that are about 25–75 mm wide and 25–300 mm long. They typically contain around 200 eggs or so on average (Dodd 2013). The freshly laid ova are black or brownish above and whitish below, and each is surrounded a single gelatinous envelope that is 3.8–5.6 mm in diameter. Females are capable of laying several thousand eggs per year. Reported clutch sizes include 800–2,000 eggs in Connecticut specimens (Ball 1936), 3,522–4,469 ova (mean 3,838) in three gravid females from Arkansas (Trauth et al. 1990), 1177-5395 ova (mean = 2,518) for 10 specimens from Virginia (Mitchell and Pague 2014), and 3,078–5,468 eggs (mean = 3,872) for 11 females from Illinois (Palis; AmphibiaWeb 2022).
The embryos can sometimes hatch in a day if breeding occurs during the warmest months of the year, but may require as long as 15 days depending on water temperatures and the passage of cold fronts (Dodd 2013). Eggs that were observed over a 4-year span by Richmond (1947) in Virginia required from 1-7 days to hatch, and those that experienced cold temperatures in March suffered very high mortality. Gosner and Black (1955) also showed experimentally that prolonged exposure to temperatures < 4–6°C will kill the developing embryos.
Aquatic Life History:
The embryos hatch at premature stages and spend the first few days attached with their ventral suckers to the jelly masses or vegetation. During this period the mouthparts, operculum, tail fins and other body parts develop. The hatchlings soon become free swimming tadpoles. For a few days they swim through the water column while remaining randomly dispersed and filter-feed on phytoplankton. During this period the mandibles and cornified teeth complete their development. The tadpoles then shift to becoming omnivores and consume a variety of food items, including periphyton, phytoplankton, zooplankton, decaying plant material, and dead animal remains. They will also eat the eggs and small larvae of both their own species and other anurans that share a breeding site. The tadpoles very likely kill and consume numerous benthic invertebrates such as midge larvae and other aquatic insects (Petranka and Kennedy 1999, Richmond 1947), but this has not been fully documented.
As larvae mature they shift to forming dense feeding aggregates in shallow water, and then later stream about in large schools. A single feeding group often contains many thousands of individuals that are tightly packed and move slowly throughout the pools (Ball 1936, Richmond 1947). Moving in dense aggregations facilitates filter feeding by churning up sediments, but may also function to reduce the risk of predation or create a thermal sink that increases larval growth rates (Dodd 2013). The eggs are frequently laid in very shallow pools that have few predators, but are susceptible to pond drying. Ball (1936) found that larvae resorted to cannibalism when the tadpoles were crowded in a drying pool. However, Richmond (1947) found no evidence of cannibalism in his studies in Virginia.
The length of the larval period depends on water temperatures and crowding effects. Metamorphosis can begin in as little as 2-3 weeks after hatching or require as long as 7-8 weeks or more. Ball (1936) reported that metamorphosis required 23-40 days in Connecticut, while Richmond (1947) estimated 26 -50 days for different ponds in Virginia. Most of the variation in length of the larval period was due to differences in seasonal temperatures. Populations in Florida appear to require less than a month (Greenberg and Tanner 2004).
Richmond (1947) noted that the tadpoles stop schooling and aggregating as metamorphosis nears. Because of explosive breeding by this species, the emergence of juveniles from the ponds is highly synchronized. A series of 63 recently transformed young that Duellman and Schwartz (1958) measured from southern Florida average slightly more than 13 mm SUV, while metamorphs that were reared by Gomez-Mestre and Buchholz (2006) varied from 16-25 mm SUL at metamorphosis.
Studies of crowding effects in laboratory tanks or artificial pond communities (Semlitsch and Caldwell 1982, Wilbur et al. 1983) show that crowding acts to slow growth, retard development, reduce survivorship, and reduce the size at metamorphosis. These effects likely interact with pond hydroperiods in nature to affect reproductive success in any given year. Crowding related to the formation of feeding aggregates and schools appears to be adaptive and enhances larval growth, but crowding that occurs in contracting ponds likely has adverse consequences, including occasional cannibalism.
Terrestrial Life History:
Successful reproduction in this species tends to be episodic due to the use of shallow, ephemeral ponds. In some years no metamorphs are produced because of premature pond drying, while in northern populations eggs that are laid early in the season may suffer complete losses from exposure to cold temperatures. At the other extreme, vast numbers of juveniles may exit the ponds, particularly after tropical storms and hurricanes trigger mass breeding and allow ponds to hold water long enough for the tadpoles to metamorphose. Atkinson et al. (2021), for example, trapped over 310,000 metamorphs that were leaving a seasonal pond in Georgia one year, but relatively few in other years. Duellman and Schwartz (1958) estimated that there were 60,000 adults breeding in a large pond in southern Florida that stretched for nearly a mile. This large gathering of adults occurred after torrential rains and the number of metamorphs produced must have been enormous.
Metamorphosis is often highly synchronized in this species and the young metamorphs have been observed moving by the tens of thousands away from the breeding ponds in Florida (Dodd 2013, Neill 1957). At breeding sites in South Carolina production is often near zero in many years, with an occasional bumper crop. In years when relatively small numbers of metamorphs are produced, competition, predation, and pond hydroperiod may all contribute to low larval survival (Gibbons and Bennett 1974, Pechmann et al. 2001, Semlitsch et al. 1996).
The young metamorphs often remain near the pond margins for a week or so before moving to upland sites (Dodd 2013, Greenberg and Tanner 2004). Dispersal mostly occurs at night, although about 20% of drift fence captures by Todd and Winne (2006) at a South Carolina site occurred during the day. Pearson (1955) noted that it takes about four to six weeks for the metamorphs to reach terrestrial sites where they establish home ranges and burrow. The juveniles often move hundreds of meters from the ponds and the adults make similarly long journeys to and from the breeding sites at night. Dodd (1996) found the adults moved an average of 539 m from the breeding site (range = 95-914 m) and noted that dispersal occurs slowly, with individuals usually moving only short distances from one movement to the next. Individuals tracked by Timm et al. (2014) in Massachusetts moved a maximum distance of 3 – 449 m from a breeding pond (mean = 130 m) and tended to establish home ranges near the edges of shrubs or beneath low-growing pine branches.
The young juveniles that were studied by Pearson (1955) grew rapidly during the first several months after metamorphosing, with growth slowing thereafter. Individuals appeared to reach sexual maturity about two years after metamorphosing. Duellman and Schwartz (1958) noted that only females > 42 mm SUL were gravid in southern Florida, but Greenberg and Tanner (2005a) rarely caught individuals < 50 mm SUL at their breeding sites in north-central Florida. They suggested that the adults may not breed until 4–5 years old and appear to rarely live more than seven years. Pearson (1955) noted that 15–39% of individuals at his study sites were greater than six years old.
The subadults and adults set-up home ranges that are centered on their burrows. The burrows are dug with the spade-like structures on the hind feet and are typically about 7-31 cm deep and 2-4 cm in diameter (Dodd 2013, Pearson 1955). They are filled with loose soil that often has to be tunneled through when entering and exiting the burrow, particularly in very sandy locales. Most animals that were studied by Pearson (1955) in northern Florida rarely dug more than 18 cm deep.
Individuals typically make short excursions of a meter or less from the entrance in search of food. They are most active for the first 2-4 hours after dark when conditions are warm and moist, and sometimes before dawn (Pearson 1955, Punzo 1992a, Ryan 2015). They also may sit at the burrow entrance partially buried and ambush passing prey (Dodd 2013, Johnson 2003, Pearson 1955). Individuals appear to be generalists and feed on a wide variety of mostly ground-dwelling insects, including beetles, flies, ants, termites, grasshoppers, crickets, katydids, bugs, cicadas, aphids, and larval lepidopterans (Dodd 2013). They also take millipedes, centipedes, isopods, spiders and oligochaetes. Punzo (1992a) found the diets of
Anaxyrus terrestris
and
S. holbrookii
at a site in Florida to be very similar, with beetles, ants, spiders and termites comprising most of the food items.
Individuals often spend many days in their burrows without surfacing and are most active on warm and moist nights. Rainfall is not required to trigger surface activity (Johnson 2003, Pearson 1955). Pearson (1955) found individuals active on the ground surface on only 8% of the nights where he made detailed observations in Florida, with individuals often remaining underground for a week or more before resurfacing. In winter, individuals burrow below the frost line and can remain dormant until warm heavy rains trigger mating. Individuals often use the same burrows for prolonged periods, but sometimes construct new burrows that are typically within a few meters of the first (Johnson 2003, Pearson 1955). The home ranges in Pearson's studies (1955, 1957) averaged only 10 m2 and did not overlap within a sex. This suggests that individuals may be territorial and defend the foraging space in the immediate vicinity of their burrows. Individuals can home and are known to return to the same home range after breeding (Dodd 2013).
General Ecology
Population Ecology:
Information on the population biology of this species has been difficult to obtain because of the erratic nature of breeding and difficulty of collecting adults due to their fossorial habits. Local populations vary substantially in size from a few individuals to perhaps thousands in some areas. Duellman and Schwartz's (1958) estimate of 60,000 adults breeding at a very large breeding site in southern Florida undoubtedly is the record.
Many populations in interior regions of the range are often strongly tied to sandy alluvial areas in floodplains and are often relatively small. Reproduction is often erratic -- with the entire population sometimes skipping breeding in some years -- and larvae suffering complete mortality from prematurely pond drying in other years (Dodd 1992, 2013, Greenberg and Tanner 2005a, Palis 2012). This species appears to do well where clusters of ponds with varying hydroperiods provide an array of breeding opportunities for the adults, particularly given that reproductive failures are common (e.g., Greenberg and Tanner 2004, 2005a). The Eastern Spadefoot is relatively long-lived compared to other species such as the true toads, which likely helps to maintain populations that exploit highly ephemeral ponds with unpredictably drying patterns.
Community Ecology:
Larvae of the Eastern Spadefoot have been used in several experiments involving artificial communities (Alford 1989a, Morin 1983, Wilbur 1987, Wilbur et al. 1983). In some cases they are superior competitors to other larvae such as
Hyla chrysoscelis
and may even benefit from the presence of certain species such as
Anaxyrus americanus
and
Lithobates sphenocephalus
. Predators such as the Eastern Newt and a centrarchid fish are highly effective predators in tanks and experimental ponds and can alter the survivorship and growth trajectories of larvae.
Adverse Environmental Impacts
Status in North Carolina
NHP State Rank:
S5
Global Rank:
G5
Environmental Threats:
The loss and degradation of both aquatic and terrestrial habitats is the biggest concern. This species has been impacted by residential development where natural sandy soils are often replaced by turf grasses or other substrates that are difficult to burrow into (Delis et al. 1996). The small, seasonal wetlands that are used as breeding sites rarely have any legal protection and countless thousands have been destroyed across the range of this species.
Status Comments:
The status of the Eastern Spadefoot has proven challenging to assess because the adults are rarely observed on the ground surface and the breeding bouts often last only a day or two (Gibson and Anthony 2019). Populations in North Carolina generally appear to be stable and there is no evidence of widespread, marked declines. There is concern that populations in the northeastern portion of the Coastal Plain may be in decline (Micancin et al. 2012).
Stewardship:
Populations are best maintained by having a series of seasonal ponds of varying hydroperiods in close proximity. Sandy or otherwise friable soils are essential. This species tolerates prescribed burning well and can quickly burrow into the soil to escape approaching flames.
Photo Gallery for
Scaphiopus holbrookii
- Eastern Spadefoot
45 photos are available. Only the most recent 30 are shown.
Recorded by: R. Newman
Carteret Co.
Recorded by: Mark Shields
Onslow Co.
Recorded by: Mark Shields
Onslow Co.
Recorded by: Mark Shields
Onslow Co.
Recorded by: Mark Shields
Onslow Co.
Recorded by: Max Ramey
Wilkes Co.
Recorded by: Tiffany Huffman
Catawba Co.
Recorded by: Tiffany Huffman
Catawba Co.
Recorded by: Tiffany Huffman
Catawba Co.
Recorded by: S. Dagenhart
Dare Co.
Recorded by: B. Bockhahn
New Hanover Co.
Recorded by: B. Bockhahn
Durham Co.
Recorded by: B. Bockhahn
Dare Co.
Recorded by: Mark Basinger
Wilson Co.
Recorded by: Rob Van Epps
Mecklenburg Co.
Recorded by: Rob Van Epps
Mecklenburg Co.
Recorded by: B. Bockhahn, J. Anderson
Harnett Co.
Recorded by: D. Brown
Harnett Co.
Recorded by: Matt Windsor
Surry Co.
Recorded by: J. Fields, J. Conolly
Cumberland Co.
Recorded by: Randy Newman
Carteret Co.
Recorded by: K. Bischof
Beaufort Co.
Recorded by: K. Bischof
Beaufort Co.
Recorded by: K. Bischof
Beaufort Co.
Recorded by: K. Bischof
Beaufort Co.
Recorded by: Randy Newman
Carteret Co.
Recorded by: N. Ward
Dare Co.
Recorded by: Carla Edwards
New Hanover Co.
Recorded by: Randy Newman
Carteret Co.
Recorded by: Randy Newman
Carteret Co.

 »
»